
感觉系统/视觉颜色感知
人类(以及猴子和猩猩等灵长类动物)在哺乳动物中拥有最佳的颜色感知能力[1]。因此,颜色在各种各样的方面发挥重要作用并非巧合。例如,颜色对于区分和识别物体、表面、自然风景甚至面部非常有用[2],[3]。颜色也是非语言交流的重要工具,包括情绪的表达[4]。
几十年来,找到颜色物理特性与其感知特性之间联系一直是一个挑战。通常,这些研究采用两种不同的方法:颜色引起的行为反应(也称为心理物理学)和它引起的实际生理反应[5]。
在这里,我们将只关注后者。颜色视觉生理基础的研究在 20 世纪下半叶之前几乎一无所知,从 1950 年开始缓慢而稳定地取得进展。在许多领域取得了重要进展,特别是在受体水平。由于分子生物学方法,以前未知的关于锥体色素遗传基础的细节得以揭示。此外,越来越多的皮质区域受到视觉刺激的影响,尽管颜色感知与受体之外的波长依赖性生理活动的相关性并不容易辨别[6]。
在本章中,我们将解释颜色感知不同过程的基本原理,从眼球中的视网膜到大脑中的视觉皮层。有关解剖学细节,请参阅本华夏公益教科书的“视觉系统解剖学”部分。
人类可以识别的所有颜色都可以通过三种主要(基本)颜色的混合来产生。受这种颜色混合理念的启发,人们提出颜色由三类传感器支持,每类传感器对可见光谱的不同部分具有最大敏感度[1]。1853 年,人们首次明确提出,正常颜色匹配存在三个自由度[7]。这后来在 1886 年得到证实[8](与最近的研究结果非常接近[9],[10])。
这些提议的颜色传感器实际上就是所谓的锥体细胞(注意:在本章中,我们将只讨论锥体细胞。棒体细胞只在弱光条件下对视觉有贡献。尽管已知它们会影响颜色感知,但其影响非常小,可以忽略不计。)[11]。锥体细胞是视网膜中发现的两种光感受器细胞类型之一,其中大量集中在中央凹处。下表列出了三种类型的锥体细胞。它们通过不同的视紫红质色素类型区分。它们相应的吸收曲线显示在下图中。
| 名称 | 对颜色的更高敏感度 | 吸收曲线峰值 [nm] |
|---|---|---|
| S, SWS, B | 蓝色 | 420 |
| M, MWS, G | 绿色 | 530 |
| L, LWS, R | 红色 | 560 |
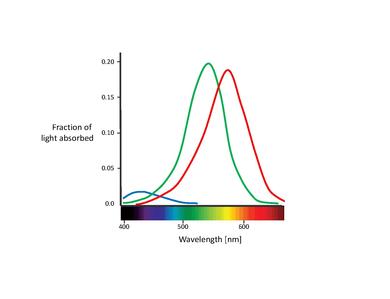
虽然对于不同锥体细胞类型的命名尚未达成共识,但最常用的名称是指它们的行动光谱峰值或它们本身对的颜色敏感度(红色、绿色、蓝色)[6]。在这篇文本中,我们将使用 S-M-L 命名法(分别代表短、中、长波长),因为这些名称更具描述性。“蓝绿红”命名法有些误导,因为所有类型的锥体细胞都对很大范围的波长敏感。
三种锥体细胞类型的一个重要特征是它们在视网膜中的相对分布。事实证明,S 锥体细胞在整个视网膜中的浓度相对较低,在中央凹的最中心区域完全不存在。实际上,它们之间的间距太大,无法在空间视觉中发挥重要作用,尽管它们能够介导弱边界感知[12]。中央凹主要由 L 锥体细胞和 M 锥体细胞组成。后两种类型的比例通常以比率衡量。L/M 比率报告了不同的值,范围从 0.67[13]到 2[14],后者是最被接受的值。L 锥体细胞几乎总是比 M 锥体细胞数量多,其原因尚不清楚。令人惊讶的是,相对锥体细胞比例几乎对颜色视觉没有显著影响。这清楚地表明,大脑具有可塑性,能够从它接收到的任何锥体细胞信号中提取意义[15],[16]。
还需要注意的是 L 锥体细胞和 M 锥体细胞吸收光谱的重叠。虽然 S 锥体细胞吸收光谱是明显分开的,但 L 锥体细胞和 M 锥体细胞峰值之间只有约 30 nm 的距离,它们的光谱曲线也存在显著的重叠。这导致这两类锥体细胞的感光量高度相关。这可以用这样一个事实来解释:为了在中央凹中心实现尽可能高的锐度,视觉系统平等地对待 L 锥体细胞和 M 锥体细胞,而不考虑它们的光谱吸收。因此,任何差异都会导致亮度信号恶化[17]。换句话说,L 锥体细胞和 M 锥体细胞光谱之间的微小分离可以解释为高对比度颜色视觉和高锐度亮度视觉需求之间的折衷。这与中央凹中心部分没有 S 锥体细胞相一致,因为中央凹中心部分的视觉锐度最高。此外,L 锥体细胞和 M 锥体细胞吸收光谱之间的紧密间距也可以用它们的遗传起源来解释。据推测,这两类锥体细胞都是从一个共同的祖先最近(大约 3500 万年前)进化而来的,而 S 锥体细胞可能在更早之前就从祖先受体中分离出来[11]。
三种不同类型锥体细胞的光谱吸收函数是人类颜色视觉的标志。该理论解决了一个长期存在的问题:虽然我们能看到数百万种不同的颜色(人类可以区分 700 万到 1000 万种不同的颜色[5]),但我们的视网膜 simply does not have enough space to accommodate an individual detector for every color at every retinal location.
从视网膜传递到更高层次的信号不是受体信号的简单逐点表示,而是受体信号的复杂组合。本节的目的是简要介绍部分信息传递的路径。
一旦视网膜上的光学图像被转换为光感受器中的化学和电信号,幅度调制信号就会在神经节细胞和更高层次转换为频率调制表示。在这些神经细胞中,信号的幅度用细胞每秒发出的电压尖峰数量表示,而不是用细胞膜上的电压差表示。为了解释和表示这些细胞的生理特性,我们会发现感受野的概念非常有用。
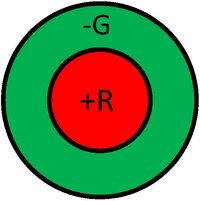
感受野是视觉区域中给定细胞响应区域的图形表示。此外,响应的性质通常在感受野的不同区域中表示出来。例如,我们可以将光感受器的感受野视为一个小的圆形区域,代表该特定受体在视觉区域的灵敏度的大小和位置。下图显示了神经节细胞的示例性感受野,通常以中心-周围拮抗的形式。图中左侧感受野说明了正中心响应(称为中心-ON)。这种响应通常是由单个锥体产生的正输入以及由几个相邻锥体产生的负响应共同生成的。因此,这种神经节细胞的响应将由来自不同锥体的正负输入组成。这样一来,细胞不仅对光点做出反应,而且充当边缘(更准确地说,是斑点)探测器。类比于计算机视觉术语,我们可以将神经节细胞的响应视为对边缘探测器核卷积后的输出。图中右侧感受野说明了负中心响应(称为中心-OFF),其可能性相同。通常,中心-ON和中心-OFF细胞将出现在同一空间位置,由相同的光感受器供给,从而导致增强的动态范围。
下图显示了除了空间拮抗之外,神经节细胞还可以具有光谱对抗性。例如,下图左侧说明了红-绿对抗响应,其中中心由来自L锥体的正输入供给,周围由来自M锥体的负输入供给。另一方面,下图右侧说明了这种细胞的中心-OFF版本。因此,在视觉信息甚至离开视网膜之前,处理就已经发生了,对颜色外观产生了深刻的影响。还有其他类型和品种的神经节细胞响应,但它们都共享这些基本概念。



在通往初级视觉皮层的路上,神经节细胞的轴突聚集起来形成视神经,它投射到丘脑的背侧膝状核(LGN)。视神经中的编码非常高效,使神经纤维的数量降至最低(受视神经大小的限制),从而也使视网膜盲点的尺寸尽可能小(约 5° 宽 x 7° 高)。此外,所呈现的神经节细胞对均匀照度将没有反应,因为正负区域是平衡的。换句话说,传输的信号是不相关的。例如,来自自然场景相邻部分的信息在空间上高度相关,因此高度可预测[18]。相邻视网膜神经节细胞之间的侧抑制最大程度地减少了这种空间相关性,从而提高了效率。我们可以将其视为在视网膜中进行的一种图像压缩过程。
鉴于L-和M-锥体吸收光谱的重叠,它们的信号也高度相关。在这种情况下,通过组合锥体信号来最大程度地减少这种相关性,可以提高编码效率。我们可以使用主成分分析 (PCA) 更容易地理解这一点。PCA 是一种统计方法,用于通过将原始变量转换为一组新的变量(主成分 (PC))来降低给定变量集的维数。第一个 PC 解释了原始变量中总方差的最大量,第二个 PC 解释了第一个成分未解释的最大方差,依此类推。此外,PC 在参数空间中线性无关且彼此正交。PCA 的主要优点是,只有几个最强的 PC 就足以覆盖系统变异性的绝大多数[19]。这种方案已应用于锥体吸收函数[20],甚至应用于自然发生的频谱[21],[22]。在锥体激发空间中发现的 PC 是 1) 亮度轴,其中 L- 和 M- 锥体信号相加 (L+M),2) L- 和 M- 锥体信号的差 (L-M),以及 3) 色彩轴,其中 S- 锥体信号与 L- 和 M- 锥体信号的总和之差 (S-(L+M))。这些从数学/计算方法导出的通道与在电生理实验中发现的三个视网膜-膝状核通道相吻合[23],[24]。使用这些机制,视觉冗余信息在视网膜中被消除。
实际上,有三个信息通道将信息从视网膜通过神经节细胞传递到 LGN。它们不仅在色度特性上不同,而且在解剖学基础上也不同。这些通道对基本颜色任务(如检测和辨别)构成重要限制。
在第一个通道中,L- 和 M- 锥体的输出协同传递到弥散双极细胞,然后传递到 LGN 的大细胞层 (M-) 中的细胞(不要与视网膜的 M- 锥体混淆)[24]。M- 细胞的感受野由中心和周围组成,它们在空间上是拮抗的。M- 细胞对亮度刺激具有高对比度敏感性,但它们在 L-M 对抗输入的某些组合下没有反应[25]。然而,由于不同 M- 细胞的零点略有不同,总体响应实际上永远不会真正为零。这种特性实际上被传递到以 M- 细胞输入为主的皮层区域[26]。
小细胞通路 (P-) 起源于 L- 或 M- 锥体到矮胖双极细胞的单个输出。它们为视网膜 P- 细胞提供输入[11]。在中央凹,P- 细胞的感受野中心由单个 L- 或 M- 锥体形成。P- 细胞感受野周围的结构仍在争论中。然而,最被接受的理论指出,周围由特定类型的锥体组成,导致亮度刺激的空间拮抗感受野[27]。小细胞层约占视网膜到 LGN 总投射的 80%[28]。
最后,最近发现的细小细胞通路 (K-) 主要携带来自 S- 锥体的信号[29]。这种类型锥体的群体投射到特殊的双极细胞,而这些双极细胞反过来为特定的小型神经节细胞提供输入。这些通常不是空间拮抗的。小型神经节细胞的轴突投射到 LGN 的薄层(紧邻小细胞层)[30]。
虽然神经节细胞确实终止于 LGN(与 LGN 细胞形成突触),但神经节细胞和 LGN 细胞之间似乎存在一对一对应关系。LGN 似乎充当信号的继电站。然而,它可能在视觉功能中发挥作用,因为皮层存在神经投射回 LGN,可以作为某种类型的切换或适应反馈机制。LGN 细胞的轴突投射到枕叶的视觉区一 (V1)。
在皮层中,来自大细胞、小细胞和细小细胞通路的投射最终到达初级视觉皮层的不同层。大细胞纤维主要支配 4Cα 层和 6 层。小细胞神经元主要投射到 4Cβ、4A 层和 6 层。细小细胞神经元终止于 1、2 和 3 层的细胞色素氧化酶 (CO-) 富集斑点[31]。
一旦进入视觉皮层,视觉信息的编码变得更加复杂。与各种光感受器的输出组合并比较以产生神经节细胞响应的方式相同,各种 LGN 细胞的输出被比较并组合起来以产生皮层响应。随着信号在皮层处理链中进一步向上推进,这个过程会重复进行,复杂程度迅速增加,以至于感受野开始失去意义。然而,一些功能和过程已在视觉皮层的特定区域中被识别和研究。
在 V1 区域(纹状皮层),双拮抗神经元(其感受野在色度和空间上都与其单个感受野的兴奋/抑制区域相反)比较视觉空间中的颜色信号[32]。它们占 V1 中细胞的 5% 到 10%。它们粗糙的大小和较小的比例与颜色视觉的空间分辨率差相匹配[1]。此外,它们对移动刺激的方向不敏感(不像其他一些 V1 神经元),因此不太可能对运动感知做出贡献[33]。然而,鉴于它们特殊的感受野结构,这类细胞是颜色对比效应的神经基础,也是编码颜色的有效手段[34],[35]。其他 V1 细胞对其他类型的刺激做出反应,例如定向边缘、各种空间和时间频率、特定空间位置以及这些特征的组合等等。此外,我们可以找到线性组合来自 LGN 细胞的输入的细胞,以及执行非线性组合的细胞。这些反应是支持高级视觉能力所必需的,例如颜色本身。

与 V1 相比,V2 中单个神经元の色度特性信息要少得多。乍一看,V1 和 V2 的颜色编码似乎没有重大差异[36]。一个例外是出现了一类新的颜色复杂细胞[37]。因此,有人认为 V2 区域参与了色调的阐述。然而,这仍然存在很大争议,尚未得到证实。
在 V1 中发现功能性眼优势后,按照模块化概念,并考虑到 P、M 和 K 通路之间的解剖学分离(在第 3 节中描述),有人提出视觉皮层中应该存在一个专门的系统来分析颜色信息[38]。V4 是历史上最受关注的可能的大脑“颜色区域”。这是因为一项有影响力的研究声称 V4 包含 100% 的色调选择性细胞[39]。然而,这一说法受到随后多项研究的质疑,有些研究甚至报告说,只有 16% 的 V4 神经元表现出色调调节[40]。目前,最被接受的概念是 V4 不仅对颜色有贡献,而且对形状感知、视觉注意力和立体视觉也有贡献。此外,最近的研究集中在其他大脑区域,试图找到大脑的“颜色区域”,例如 TEO[41] 和 PITd[42]。这些区域之间的关系仍在争论中。为了调和讨论,有些人使用术语“后颞下(PIT)皮层”来表示包括 V4、TEO 和 PITd 的区域[1]。
如果 V1、V2 和 V4 细胞中的皮层响应已经是一项非常复杂的任务,那么大约 30 个视觉区域网络中复杂视觉响应的复杂程度就非常巨大。图 4 显示了已识别的不同皮层区域(不是细胞)的连接性的一小部分[43]。
在这个阶段,用简单的术语解释单个皮层细胞的功能变得极其困难。事实上,单个细胞的功能可能没有意义,因为各种感知的表征必须分布在整个皮层中的一组细胞中。
颜色视觉适应机制
[edit | edit source]尽管研究人员一直在试图解释人类视觉系统中颜色信号的处理过程,但重要的是要理解颜色感知不是一个固定过程。实际上,有各种动态机制用于根据观看环境优化视觉响应。与颜色感知特别相关的是暗适应、亮适应和色度适应的机制。
暗适应
[edit | edit source]暗适应是指当照明水平降低时发生的视觉灵敏度变化。视觉系统对光照降低的反应是变得更加敏感,提高其即使在光照条件不佳的情况下也能产生有意义的视觉响应的能力[44]。
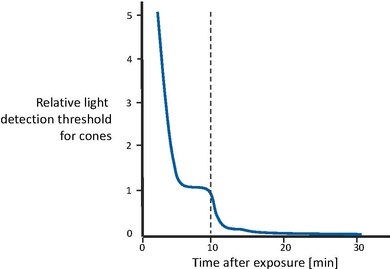
图 5 显示了从极高照明水平过渡到完全黑暗后视觉灵敏度的恢复[43]。首先,视锥细胞逐渐变得更加敏感,直到在几分钟后曲线趋于平稳。然后,在大约 10 分钟后,视觉灵敏度大致保持恒定。此时,视杆系统恢复了足够的灵敏度,其恢复时间更长,能够胜过视锥细胞,因此控制了整体灵敏度。视杆细胞灵敏度也逐渐提高,直到在大约 30 分钟后变得渐近。换句话说,视锥细胞负责最初 10 分钟的灵敏度恢复。之后,视杆细胞的性能优于视锥细胞,在大约 30 分钟后获得完全灵敏度。
这只是为了尽可能好地适应黑暗光照条件而产生的几种神经机制之一。其他一些神经机制包括著名的瞳孔反射、光色素的消耗和再生、视网膜细胞中的增益控制和其他更高级别的机制以及认知解释等等。
亮适应
[edit | edit source]亮适应本质上是暗适应的逆过程。事实上,这两个过程的基础生理机制是相同的。但是,重要的是要单独考虑它,因为它的视觉特性有所不同。

亮适应发生在照明水平提高时。因此,视觉系统必须变得不那么敏感,以便在有大量可见光可用时产生有用的感知。视觉系统用于产生感知的信号具有有限的输出动态范围。但是,现实世界中的照明水平至少覆盖了 10 个数量级的变化。幸运的是,我们很少需要同时查看整个照明范围。
在高光照水平下,适应是通过光色素漂白实现的。这会调整受体中的光子捕获,并保护视锥细胞响应在明亮背景下不被饱和。亮适应的机制主要发生在视网膜内[45]。事实上,增益变化在很大程度上是视锥细胞特异性的,并且适应会将信号池化到不超过单个视锥细胞直径的区域[46],[47]。这表明亮适应的定位可能早在受体阶段就开始了。然而,似乎存在不止一个灵敏度缩放位点。有些增益变化非常快,而另一些则需要几秒甚至几分钟才能稳定[48]。通常,亮适应需要大约 5 分钟(比暗适应快六倍)。这可能表明了受体后位点的影响。
图 6 显示了亮适应的示例[43]。如果我们使用单个响应函数将大范围的强度映射到视觉系统的输出,那么对于给定的场景,我们只能使用非常小的范围。很明显,使用这种响应函数,任何给定场景的感知对比度将受到限制,并且由于信噪比问题,对变化的视觉灵敏度会严重降低。这种情况由虚线表示。另一方面,实线代表视觉响应的曲线族。这些曲线将给定场景中的有用照明范围映射到视觉输出的完整动态范围,从而为每种情况产生最佳的视觉感知。亮适应可以看作是将视觉响应曲线沿着照明水平轴滑动,直到达到给定观看条件下的最佳水平。
色彩适应的概念通常是指三种锥体光谱响应曲线高度的变化。这种调整的出现是因为光适应在每种锥体类型中独立发生。该假设的具体公式被称为 *冯·克莱斯适应*。该假设指出,适应反应分别在三种锥体类型中发生,并且等效于将它们固定的光谱敏感度乘以一个比例常数[49]。如果比例权重(也称为 *冯·克莱斯系数*)与每种锥体类型的光吸收成反比(即,较低的光吸收需要较大的系数),那么冯·克莱斯缩放将保持每种锥体类别内的平均响应恒定。这为保持物体在照明变化的情况下感知到的颜色提供了一种简单而强大的机制。在多种不同的条件下,冯·克莱斯缩放很好地解释了光适应对颜色敏感度和外观的影响[50],[51]。
观察不同照明条件下白色物体是最容易理解色彩适应的方式。例如,考虑在日光、荧光灯和白炽灯下观察一张纸。日光含有比荧光灯更多的短波能量,而白炽灯含有比荧光灯更多的长波能量。然而,尽管照明条件不同,纸张在三种光源下都近似保持其白色外观。这是因为 S 锥体系统在日光下变得相对不那么敏感(为了补偿额外的短波能量),而 L 锥体系统在白炽灯下变得相对不那么敏感(为了补偿额外的长波能量)[43]。
- ↑ a b c d Conway, Bevil R (2009). "颜色视觉、锥体和皮层中的颜色编码". 神经科学家. 15: 274–290.
- ↑ Russell, Richard and Sinha, Pawan} (2007). "现实世界中的面部识别:表面反射属性的重要性". 知觉. 36 (9).
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Gegenfurtner, Karl R and Rieger, Jochem (2000). "颜色对自然场景识别的感官和认知贡献". 当代生物学. 10 (13): 805–808.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Changizi, Mark A and Zhang, Qiong and Shimojo, Shinsuke (2006). "裸露的皮肤、血液和灵长类颜色视觉的进化". 生物学快报. 2 (2): 217–221.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ a b Beretta, Giordano (2000). 理解颜色. 惠普公司.
- ↑ a b Boynton, Robert M (1988). "颜色视觉". 心理学年度回顾. 39 (1): 69–100.
- ↑ Grassmann, Hermann (1853). "关于颜色混合理论". 物理学年鉴. 165 (5): 69–84.
- ↑ Konig, Arthur and Dieterici, Conrad (1886). "基本感觉及其在光谱中的强度分布". 普鲁士皇家科学院.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Smith, Vivianne C and Pokorny, Joel (1975). "中央凹锥体光感受器在 400 到 500 纳米之间的光谱敏感度". 视觉研究. 15 (2): 161–171.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Vos, JJ 和 Walraven, PL (1971)。“关于中央凹视锥细胞原色的推导”。视觉研究。11 (8): 799–818.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ a b c Gegenfurtner, Karl R 和 Kiper, Daniel C (2003)。“色觉”。神经科学。26 (1): 181.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Kaiser, Peter K 和 Boynton, Robert M (1985)。“蓝色机制在波长辨别中的作用”。视觉研究。125 (4): 523–529.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Paulus, Walter 和 Kroger-Paulus, Angelika (1983)。“视网膜颜色编码的新概念”。视觉研究。23 (5): 529–540.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Nerger, Janice L 和 Cicerone, Carol M (1992)。“人类旁中央凹视网膜中 L 视锥细胞与 M 视锥细胞的比例”。视觉研究。32 (5): 879–888.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Neitz, Jay 和 Carroll, Joseph 和 Yamauchi, Yasuki 和 Neitz, Maureen 和 Williams, David R (2002)。“色觉由一种可塑的神经机制介导,这种机制在成年人中是可调节的”。神经元。35 (4): 783–792.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Jacobs, Gerald H 和 Williams, Gary A 和 Cahill, Hugh 和 Nathans, Jeremy (2007)。“在表达人类视锥细胞光色素的转基因小鼠中出现的新的色觉”。科学。315 (5819): 1723–1725.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Osorio, D 和 Ruderman, DL 和 Cronin, TW (1998)。“估计灵长类动物视网膜编码的亮度信号中由红视锥细胞和绿视锥细胞对自然图像进行采样而产生的误差”。JOSA A。15 (1): 16–22.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Kersten, Daniel (1987)。“自然图像的可预测性和冗余”。JOSA A。4 (112): 2395–2400.
- ↑ Jolliffe, I. T. (2002)。主成分分析。施普林格出版社.
- ↑ Buchsbaum, Gershon 和 Gottschalk, A (1983)。“三色视觉、对立色编码和视网膜中的最佳颜色信息传输”。伦敦皇家学会会刊。B辑。生物科学。220 (1218): 89–113.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Zaidi, Qasim (1997)。“L 视锥细胞和 M 视锥细胞信号的去相关”。JOSA A。14 (12): 3430–3431.
- ↑ Ruderman, Daniel L 和 Cronin, Thomas W 和 Chiao, Chuan-Chin (1998)。“视锥细胞对自然图像的响应统计:对视觉编码的意义”。JOSA A。15 (8): 2036–2045.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Lee, BB 和 Martin, PR 和 Valberg, A (1998). "异色闪烁光度计的生理基础,在猕猴视网膜神经节细胞中得到证实". 生理学杂志. 404 (1): 323–347.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ a b Derrington, Andrew M 和 Krauskopf, John 和 Lennie, Peter (1984). "猕猴外侧膝状体核中的色觉机制". 生理学杂志. 357 (1): 241–265.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Shapley, Robert (1990). "视觉敏感度和并行的视网膜皮层通路". 心理学年度评论. 41 (1): 635--658.
- ↑ Dobkins, Karen R 和 Thiele, Alex 和 Albright, Thomas D (2000). "人类和猕猴红绿等亮度点的比较:不同物种间 L:M 视锥比例的证据". JOSA A. 17 (3): 545–556.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Martin, Paul R 和 Lee, Barry B 和 White, Andrew JR 和 Solomon, Samuel G 和 Ruttiger, Lukas (2001). "灵长类动物外周视网膜神经节细胞的色觉敏感性". 自然. 410 (6831): 933–936.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Perry, VH 和 Oehler, R 和 Cowey, A (1984). "投射到猕猴背侧外侧膝状体核的视网膜神经节细胞". 神经科学. 12 (4): 1101--1123.
{{cite journal}}: 引文格式有误:未知参数忽略:|month=(help)CS1 maint: 多个名字:作者列表 (link) - ↑ Casagrande, VA (1994). "通往灵长类动物 V1 区的第三条平行视觉通路". 神经科学趋势. 17 (7): 305–310.
- ↑ Hendry, Stewart HC 和 Reid, R Clay (2000). "灵长类视觉中的锥体通路". 神经科学年度评论. 23 (1): 127–153.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Callaway, Edward M (1998). "猕猴初级视觉皮层的局部回路". 神经科学年度评论. 21 (1): 47–74.
- ↑ Conway, Bevil R (2001). "清醒猕猴初级视觉皮层 (V-1) 中色觉细胞的视锥输入的空间结构". 神经科学杂志. 21 (8): 2768–2783.
{{cite journal}}: 引文格式有误:未知参数忽略:|month=(help) - ↑ Horwitz, Gregory D 和 Albright, Thomas D (2005). "猕猴 V1 中色觉线性运动检测器的稀缺". 视觉杂志. 5 (6).
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Danilova, Marina V 和 Mollon, JD (2006). "空间分离颜色的比较". 视觉研究. 46 (6): 823–836.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Wachtler, Thomas 和 Sejnowski, Terrence J 和 Albright, Thomas D (2003). "清醒猕猴初级视觉皮层中色觉刺激的表征". 神经元. 37 (4): 681–691.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Solomon, Samuel G 和 Lennie, Peter (2005). "视觉皮层神经元中的色觉增益控制". 神经科学杂志. 25 (19): 4779–4792.
{{cite journal}}: CS1 maint: 多个名字:作者列表 (link) - ↑ Hubel, David H (1995). 眼、脑和视觉. 科学美国人图书馆/科学美国人书籍.
- ↑ Livingstone, Margaret S 和 Hubel, David H (1987). “心理物理学证据表明,感知形状、颜色、运动和深度的通道是分开的”。神经科学杂志. 7 (11): 3416–3468.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ Zeki, Semir M (1973). “猕猴前纹状皮层的颜色编码”。脑研究. 53 (2): 422–427.
- ↑ Conway, Bevil R 和 Tsao, Doris Y (2006). “fMRI 揭示清醒猕猴皮层的颜色结构”。大脑皮层. 16 (11): 1604–1613.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ Tootell, Roger BH、Nelissen, Koen、Vanduffel, Wim 和 Orban, Guy A (2004)。“在猕猴视觉皮层中寻找颜色‘中心’”。大脑皮层. 14 (4): 353–363.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ Conway, Bevil R、Moeller, Sebastian 和 Tsao, Doris Y (2007)。“猕猴外纹状皮层的特化颜色模块”。560--573. 56 (3): 560–573.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ a b c d Fairchild, Mark D (2013)。颜色外观模型. 约翰·威利父子公司.
- ↑ Webster, Michael A (1996)。“人类颜色感知及其适应”。网络:神经系统中的计算. 7 (4): 587–634.
- ↑ Shapley, Robert 和 Enroth-Cugell, Christina (1984)。“视觉适应和视网膜增益控制”。视网膜研究进展. 3: 263–346.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ Chaparro, A、Stromeyer III, CF、Chen, G 和 Kronauer, RE (1995)。“人类视锥细胞似乎在低光照条件下会适应:对红绿检测机制的测量”。视觉研究. 35 (22): 3103–3118.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ Macleod, Donald IA、Williams, David R 和 Makous, Walter (1992)。“由单个视锥细胞馈送的视觉非线性”。视觉研究. 32 (2): 347–363.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ Hayhoe, Mary (1991)。颜色和亮度的适应机制. 春季.
- ↑ MacAdam, DAvid L (1970)。颜色科学的来源. 麻省理工学院出版社.
- ↑ Webster, Michael A 和 Mollon, JD (1995)。“对比度适应影响的颜色恒常性”。自然. 373 (6516): 694–698.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link) - ↑ Brainard, David H 和 Wandell, Brian A (1992)。“不对称颜色匹配:颜色外观如何依赖于光源”。JOSA A. 9 (9): 1443–1448.
{{cite journal}}: CS1 maint: 多个名称:作者列表 (link)