
结构生物化学/衰老与代谢控制分析
衰老是指机体(如细胞)在其一生中积累了过量的损伤。由此产生的损伤最终会影响机体的整体生存和状态。随着机体衰老,“其输出的退化导致功能下降和死亡,这是衰老的结果”(1)。这种输出的退化与细胞的代谢历史有关,而代谢历史会影响细胞的功能。
衰老的概念导致人们普遍认为,衰老是由于机体在其一生中积累的损伤导致其无法保护、维持和修复自身。科学家正在进行实验,试图确定与衰老相关的哪些损伤会导致机体功能丧失。这很困难,因为与损伤相关的许多因素在不同模型和个体之间可能会有所不同,例如损伤程度、组织类型、年龄,以及所研究的机体类型。首先,机体要生存需要做些什么?首先,机体必须能够找到食物、住所,并抵御感染或掠食者。机体必须能够避免死亡。即使消除了这些因素,死亡和功能丧失也会随着年龄的增长而发生。然而,并没有进化出导致死亡的基因。衰老的机体被认为会降低个体对下一代的遗传贡献。换句话说,这是不利的。
在 Murphy 的论文“衰老的控制理论”中,他指出“基因组及其表达方式限制了死亡率和寿命”。然而,这种观点仍然存在问题,因为所有机体的寿命各不相同,即使它们的基因非常相似。可以得出结论,导致死亡的年龄的确是遗传决定、环境变化以及机体一生中发生的各种事件的综合结果,而这些因素会对其产生影响。由于一生中低剂量暴露于紫外线辐射和伽马射线辐射等外部因素,机体执行稳态机制的能力开始下降,衰老变得明显。人们还提出,衰老反映了细胞衰老,即细胞自我复制和生长的能力不可逆转地停止。
一个障碍是突变和环境干预,这些干预阻碍了许多功能,这使得很难确定衰老的原因。如前所述,机体死亡的原因可能有很多。重要的是要看到,必须考察所有可能的生化和生理实体,并独立观察每个实体。这将有助于缩小导致衰老的原因范围。
层次结构的最高级别是机体的功能和一组参与相互作用以及与周围环境相互作用的生理系统。每个系统都以某种方式通过输入和输出与其他系统以及环境相互作用。

在上图中,为了简便起见,相互作用的生理系统被显示为两个独立的系统。它们受到来自环境和来自其他系统的输入的影响。例如,随着机体衰老,其各个系统的输出随着时间的推移而减少,机体的死亡率也会增加。对机体的损伤也会导致功能障碍,从而导致系统输出不当。
一般来说,机体内的系统通常会随着衰老而衰退。然而,很明显,不同物种的死亡率并不相同。一种理论认为,自然选择可能会影响机体的衰老过程。幸存下来的机体世代可以表现出可以通过进化保留的特征。这暗示着不同机体(从果蝇、小鼠、酵母和蠕虫)可能存在类似的衰老过程。然而,生物年龄并不可靠,因为这在不同物种之间也会有所不同。衰老的其他方面可能是器官特异性的,并且会表现为皮肤衰老的出现。对于一个独立的系统,很难确定是缺陷输入的影响还是该系统的内在损伤导致了细胞功能障碍。所有上述原因解释了为什么科学家难以确定是所有生理系统的衰退还是仅仅一个系统的衰退导致了机体衰老并最终死亡。
细胞会发生许多变化,但影响生理系统功能输出的变化会影响衰老。系统的功能障碍是由于细胞数量及其输出的减少造成的。根据 Murphy 的说法,“细胞的变化是由其代谢历史引起的,并且是由于非特异性损伤以及信号通路和基因表达的变化造成的。这些反过来会对细胞功能和细胞数量产生影响。”随着机体衰老,组织会发生许多变化。衰老会导致细胞数量发生变化。细胞数量的减少会导致维持细胞的机制出现紊乱。
一些有丝分裂组织可以从其他分化细胞中补充失去的细胞。但随着机体衰老,哺乳动物干细胞补充失去细胞的效率会降低。为了确定生理系统是否因细胞数量的减少而受损,记录系统受损之前和之后发生的细胞丢失或增益至关重要。细胞功能下降的一个例子包括神经元中的突触传递和肌肉骨骼运动单位的收缩。在哺乳动物中,研究表明,随着年龄的增长,由于突触连接和传导的减少,运动和神经功能会下降。
与限制饮食的动物有关的寿命成为机体对食物供应的反应,以及它们对环境中营养变化的反应。这些变化在细胞水平上通过能量激活来衡量。喂养模式和觅食行为的变化也是其适应阶段的指标。这些营养物质感应通路也为机体提供了时间来检测和应对资源供应的变化。然而,由于这种寿命依赖于 daf-16,这意味着这些神经元的营养物质感应并非必需的。
现在我们来看看单个细胞,细胞功能障碍和死亡是细胞代谢历史的属性。这些属性会影响系统功能,从而影响机体的死亡率。细胞的初始状态取决于其基因组,其基因组会产生发育史、机体中占据的物理生态位以及影响基因组表达的表观遗传因素。
细胞的代谢历史会导致 DNA 序列发生变化,而这些变化反过来会影响基因表达。这也会影响细胞在其一生中的增殖、功能障碍和死亡。
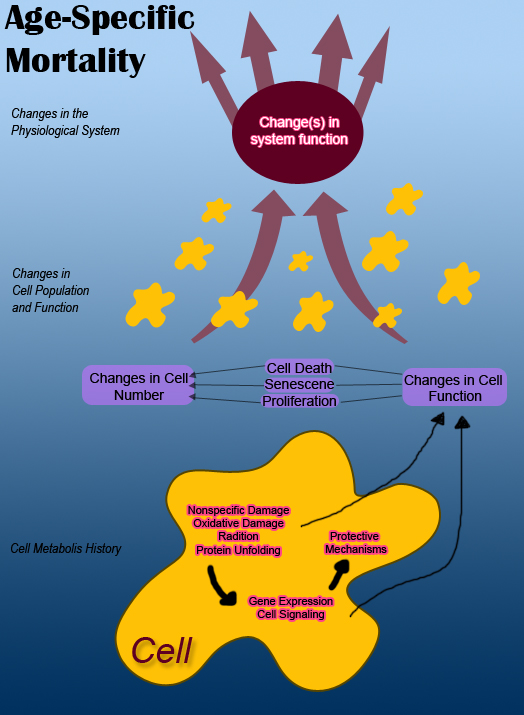

所有这些主题相互关联,也与衰老过程相关。由于细胞数量及其功能的变化,系统会发生功能障碍。细胞数量和功能的变化是由细胞的代谢历史引起的,主要涉及非特异性损伤和基因表达的变化。这些代谢历史变化的结果导致细胞数量和功能的变化。影响衰老的细胞代谢历史的重要因素和贡献因素包括非特异性损伤以及基因表达和信号通路的变化。
非特异性损伤
[edit | edit source]对生物体造成的不同类型的非特异性损伤会导致细胞功能丧失和功能障碍。一些损伤包括氧化损伤、辐射或化学反应。随着时间的推移,非特异性损伤最终会损伤和阻碍分子。细胞尽力生存并保护自己免受损伤。在能够修复错误折叠的蛋白质的情况下,细胞会启动损伤修复,但诸如 DNA 突变的修复之类的损伤无法解决。这种也是不可逆的非特异性损伤会严重损害细胞及其功能。随之而来的问题是,对细胞造成的损伤是否会对系统的输出产生足够的影响以导致衰老。
基因表达和细胞信号传导
[edit | edit source]关于非特异性损伤,它会影响细胞的基因表达和细胞信号传导。受非特异性损伤影响的基因表达也会影响其对系统输出的影响,从而影响正常衰老过程中的死亡率。有证据表明,在正常衰老过程中会发生大量的基因表达变化,但它们对衰老过程的贡献会因系统而异,总体而言也会因生物体而异。
代谢控制分析与衰老
[edit | edit source]代谢控制分析 (MCA) - 一种实验方法,用于了解控制在代谢途径和网络中是如何分布的。
为了确定与衰老过程相关的因素,必须对所分析的系统和观察到的可测量变量进行设定限制。一旦这些因素被定义和限制,就可以测试和操纵所讨论的生理系统,以便可以看到该变量与其控制的步骤之间的关系。在 MCA 实验期间,该变量的控制程度也将被确定。通过使用 MCA,已经开发出强大的数学形式化方法来观察整个系统,以及以下内容:定量确定控制程度、揭示所有控制部位以及控制步骤的影响。虽然已经表明,生化途径和机制的极端和较大变化会增加或减少寿命,但没有足够的证据表明它们会影响正常衰老。与代谢途径和衰老进行比较,MCA 可以告诉人们有关控制系统中特定过程的步骤的重要信息。
MCA 在衰老中的应用
[edit | edit source]MCA 过程及其在系统中的应用需要以某种方式进行才能获得结果。由于主题是生物体的衰老,因此需要与机体衰老相关的死亡率读数。如前一节所述,可以对系统因素进行微小改变,并通过利用数学方法获得可量化的结果。需要测量死亡率读数以响应系统中所做的微小变化。请记住,因素的变化是由于认为它可能控制衰老。为了使一个因素影响衰老,它必须改变相应的死亡率和对衰老的贡献。MCA 实验是在从果蝇到小鼠的各种动物种群中进行的。然后可以测量死亡率读数并进行比较,以查看物种之间是否存在相似性。
在上图中,死亡率读数通过将其绘制在与衰老相关的变量(已增加和减少)上显示。深蓝色阴影区域表示变量在正常衰老过程中是如何改变的。该图还显示了正常衰老的不同情况。变量的影响可以降低或增加,这也影响死亡率。这是一个在 MCA 实验后可以绘制的图表示例,以展示一个人的发现和结果。主要思想是,如果选择影响正常衰老的正确变量,死亡率曲线将具有可测量的斜率。
染色质在衰老中的作用
[edit | edit source]染色质 首先是在胚胎发生过程中在生物体中形成的。这种遗传物质易于通过 DNA 甲基化和不同类型的组蛋白修饰进行修饰;这些变化不会影响 DNA 的核苷酸序列。这些变化在生物体一生中的积累导致随机、随机的结构。在一项对丹麦双胞胎的实验中,观察到亲属之间的变异是染色质变化导致的遗传转移的结果。这支持了这样一个观点,即一种理论引用了对丹麦双胞胎的实验,并提出衰老是染色质结构变化积累的结果(2)。有几种与染色质修饰相关的蛋白质。这些蛋白质直接影响转录调控,但也影响染色质中基因的结构和表达。
CDKN2a (p16INK4a)
[edit | edit source]CDKN2a 通过阻止 CDK4/CDK6 磷酸化视网膜母细胞瘤蛋白来调节视网膜母细胞瘤蛋白,视网膜母细胞瘤蛋白抑制肿瘤。CDKN2a 表达增加与细胞衰老和衰老相关,这与细胞压力水平升高有关。由于这些关联,已经建议使用这种蛋白质作为衰老的生物标志物。在一项对小鼠 BubR1 半不足的实验中,发现消除 CDKN2a 阳性细胞的受试者寿命延长,并且与年龄相关的退化减少。实验还表明,在老年小鼠中去除 CDKN2a 阳性细胞改善了由于衰老引起的退化,并延长了这些影响。CDKN2a 表达受三胸 (TrxG) 和多梳 (PcG) 控制,它们分别打开和关闭基因。
SIRT1 和 SIRT6
[edit | edit source]SIRT1 和 SIRT6 都是 NAD+ 依赖性赖氨酸脱乙酰基酶,在 DNA 修复和衰老中发挥作用。已在酵母中发现并对 SIRT1 和 SIRT6 的同源蛋白 Sir2 进行了实验。这种蛋白质使 rRNA 和端粒沉默。如果没有该基因的表达,那么 rDNA 阵列将从基因组中去除以形成环状 DNA,并且酵母的寿命会下降。类似地,SIRT1 和 SIRT6 会影响细胞对 DNA 修复和压力水平的反应。这两种蛋白质通过 DNA 修复过程和转录调控影响衰老过程。
当 DNA 发生氧化损伤时,SIRT1 会分散到所有修复位点,在那里 H1K26 被脱乙酰化以增加 DNA 修复。然而,当 SIRT1 被招募到基因组的受损区域时,需要共济失调毛细血管扩张突变途径激活,它会脱乙酰化 Nijmegen 断裂综合征蛋白 (NBS1),NBS1 控制 DNA 修复途径,但也控制损伤依赖性转录失调。因此,虽然 SIRT1 维持 DNA,但它在衰老中也起着至关重要的作用。SIRT1 表达增加不会延长生物体的寿命,但会增加其肿瘤抵抗力,改善其代谢,并预防糖尿病。SIRT1 还起着刺激非组蛋白的作用,例如 p53 (肿瘤抑制基因)、FOXO(转录框)和 Ku70(DNA 修复因子)。
然而,SIRT6 的过度表达确实会增加小鼠的寿命(尤其是在雄性小鼠中)。SIRT6 是一种调节蛋白,通过附着在活化 B 细胞核因子 κ 轻链增强子(NF-kB)的活性位点并抑制其转录来影响其依赖性炎症反应。因此,SIRT6 表达降低会导致 NF-kB 反应增加。缺乏 SIRT6 的人类更容易受到攻击 DNA 的药物的攻击。SIRT6 还参与同源重组,调节对 DNA 损伤的反应,并通过增加端粒功能障碍来促进衰老。
总体而言,MCA 方法的开发极大地提高了我们对影响和影响正常衰老过程的因素的理解。MCA 方法应与测试衰老的实验模型一起应用,因为它将使我们能够解决可能影响衰老的因素。
- Murphy, Michael P. and Partridge, Linda. “Toward a Control Theory of Aging.” ["http://www.ncbi.nlm.nih.gov/pubmed/18318658"], 'Annual Review of Biochemistry', 2008.
- http://www.annualreviews.org/doi/pdf/10.1146/annurev.biochem.77.061206.171059
- O'Sullivan, Roderick and Jan Karlseder. "The Great Unravelling: Chromatin as a Modulator of the Aging Process". ("http://www.sciencedirect.com/science/article/pii/S0968000412001132"), 'Trends in Biochemical Sciences', 1 November 2012 (Vol. 37, Issue 11, pp. 466-476)