
结构生物化学/冷适应酶
生活在永久寒冷环境中的生物体拥有在寒冷环境中高效发挥作用的酶。一些例子包括鱼类,它们也进化出抗冻蛋白作为一种适应方式,以及一些必须生活在寒冷地区的原核生物。最近的发现表明,对这些酶的进一步研究实际上可能具有生物技术应用。最常提到的冷适应酶是来自盐生伪单胞菌(AHA)的α-淀粉酶,一种原核生物。生物体在寒冷环境中茁壮成长的能力来自于其合成冷适应酶的能力。在寒冷环境中繁荣的生物被称为嗜冷菌。这些酶已经进化出一系列结构特征,使其具有高灵活性,尤其是在活性位点周围,低活化焓,低底物亲和力和低温下的高比活性。冷适应酶的结构、功能和稳定性的研究在蛋白质折叠和催化研究中处于起步阶段,这是一个仍在发展中的领域。
不同的生物体以不同的方式进化,导致它们适应不同的适合自身的热环境。极端环境下的热适应特别难以适应,并且单个生物体所能耐受的范围有限。嗜冷微生物,即适应寒冷的微生物,据说能够在-20°C的雪和冰中进行代谢。一些嗜冷菌甚至可以在≤0°C下增殖,但受限于<30°C的温度(2-5)。
由于世界上许多海洋、寒冷的高山地区、洞穴、高层大气和极地地区都发现了大量的微生物,因此地球上很大一部分生物量,或某个区域的活生物体,是在这些寒冷温度下在其特定区域产生的。来自生命三个域的生物:细菌、古菌和真核生物,也来自这些寒冷的环境。大多数冷适应酶被发现来自原核生物。为了生存,所有这些生物体都必须与其周围环境处于热平衡状态。这来自于其细胞的所有组分都适当地适应了寒冷。为了适应寒冷,已经采用了潜在的机械多样性和细胞特异性适应策略。从对这些生物体的研究中出现的一个普遍模式是,生活在永久寒冷环境中的生物体通常会进化出有助于它们在寒冷环境中发挥作用的酶。

一个将被进一步研究和解释的冷适应酶的例子是来自盐生伪单胞菌(AHA)的α-淀粉酶。这种酶是被研究最多的冷适应酶。
下面的阿伦尼乌斯方程描述了所有反应(包括酶促反应)的速率。
kcat = AKe-Ea/RT
kcat = 酶反应速率(随着绝对温度(T)的升高而增加,随着活化能(Ea)的降低而降低)
A = 指前因子
K = 动态传递系数(通常假定为1)
R = 普适气体常数(8.314 J mol−1 K−1)
根据该方程,由于0°–4°C的低温,系统可用于克服障碍的动能不足。一些有助于补偿缓慢代谢速率的策略包括:增加酶浓度的能量消耗策略,鱼类和线虫中同工酶的季节性表达,以及酶的进化,其中反应速率趋于变得与温度无关,而是接近扩散控制。冷适应酶倾向于将其最佳活性温度转移到较低的温度,同时稳定性降低。这些酶在降低基态和过渡态之间的活化自由能垒时表现出高反应速率,如下式所示
ΔG#=ΔH# -TΔS#
ΔG# = 活化自由能垒
ΔH# = 活化焓的变化
ΔS# = 活化熵的变化
T = 绝对温度
冷适应酶的结构具有高度的灵活性,这使得它们在低至中等温度下解折叠。研究人员尝试了许多方法来研究解折叠和折叠转变,以确定它们解折叠的方式并测量酶的动力学和构象稳定性。这些方法包括分光光度法、量热法和电泳法。已经使用这些技术对多结构域蛋白质(如壳寡糖酶)进行了实验。蛋白质有解折叠的趋势,并且这些过程是不可逆的。当大的多结构域蛋白质因热而解折叠时,这些蛋白质倾向于以动力学驱动的形式完成解折叠过程。
-
 壳寡糖酶的结构
壳寡糖酶的结构

AHA是唯一一个具有可逆解折叠的冷适应酶的例子。在20°C或更高的温度下,AHA具有可逆解折叠,这可以通过在初始热变性后的第二次扫描中100%恢复ΔH cal.(解折叠过程中吸收的总热量)来证明。小分子量酶通过协同解折叠过程解折叠。协同解折叠发生是因为其紧密堆积的体积导致结构元素之间相互作用的数量很少,这有助于它保持其自然状态。如果协同解折叠中数量有限的相互作用受到干扰,则由于相互作用的增加,可能会发生两态解折叠。
在胰腺猪α-淀粉酶(PPA)和更稳定的AHA突变体中,不会发生两态解折叠过程。相反,其他类型的折叠可能来自离子相互作用的增加,导致折叠过程中分子内差异的频率发生。AHA突变体显示了这种差异或发散的概念,导致热失活速率与可逆程度直接相关。
在TUG-GE酶中,在3°C和12°C的温度下,AHA可逆地解折叠并显示出两个转变。在较低尿素浓度下解折叠的转变属于活性位点区域。活性位点是由结构的协同解折叠形成的,显示了蛋白质其他更稳定区域或结构域的独立解折叠。当解折叠开始时,发现冷适应酶的底物结合区域是最灵活的区域,因为其高Km。对AHA的研究表明,活性位点区域的不稳定性是热不稳定酶的关键,并且是检查更广泛的冷适应酶的重要概念。
酶失活或变性的过程用动力学稳定性表示。大多数冷适应酶在50°C的温度下半衰期小于20分钟,有些酶甚至在较低的温度下变性。为了预测动力学稳定性,必须考虑折叠或活性状态与过渡状态之间的自由能变化的大小,如下所示
F K¦↔ 〖TS〗^# k¦→ D
F = 折叠酶
K = 平衡常数
K= 一级速率常数
D = 变性状态
冷适应酶通过降低ΔG#来提高热解折叠速率,如下式所示
ΔG#= =RT ln K
冷适应酶的热稳定性降低可能是由于折叠形式的ΔH#较低。需要断裂一定数量的一价相互作用才能达到过渡态(TS#),并且此数量的减少导致折叠形式的数量减少。谷氨酸脱氢酶是一种热稳定性低的酶的例子,这是过渡态无序性增加的直接结果。在解折叠过程中,热稳定酶熵降低的关键原因可能是非极性基团的水合作用。这是由水围绕疏水侧链形成有序结构引起的,从而降低了系统的熵。确定变性的活化参数对于发现更大范围的冷适应酶及其热稳定同源物至关重要。
通过将冷适应酶的X射线结构与来自中温生物和嗜热生物的蛋白质同源模型进行比较,已经发现X射线结构揭示了冷适应的结构基础。酶的活性与稳定性与其生理环境密切相关。研究还表明,主要利用海洋生物来研究冷适应酶。已经确定了柠檬酸合酶的X射线结构,并将其与细菌、真核生物和古菌的其他酶进行了比较。由于缺乏晶体结构信息,研究通常选择密切相关的生物体。通过最大化比较数据集中的热力学差异并最小化系统发育差异,可以弥补酶X射线结构的不足。
疏水残基与溶剂水分子之间的疏水相互作用在促进冷适应蛋白质的结构灵活性和热稳定性方面发挥着关键作用。
冷适应酶的氨基酸往往比中温生物和嗜热生物的同源物更小且疏水性更低。由于范德华力较弱、作用范围短且对距离敏感,蛋白质内部疏水基团之间的距离将决定对稳定化的焓贡献。因此,范德华力的减少和内部基团运动的增加将导致冷适应酶的不稳定。蛋白质内部的一个基团例子是异亮氨酸。由于其支链和大小,异亮氨酸可以有效地填充核心并稳定蛋白质。在冷适应的胰蛋白酶、柠檬酸合酶和AHA中,核心内异亮氨酸残基的数量更少。
疏水相互作用在室温下最强,因为疏水侧链在水中的溶解度在20°C的最低温度下达到最小。通过对31种可逆展开蛋白质的研究,发现约75%的蛋白质在室温附近具有最大稳定性。这表明蛋白质核心中的疏水相互作用在增强蛋白质在低温到中等温度下的稳定性方面发挥着关键作用。
许多酶的X射线结构研究表明,冷适应酶的表面存在较高比例的疏水或非极性残基。使用大规模建模和结构研究,发现了类似的结论,这些结论表明酶的可溶剂化表面或埋藏表面的平均分数在冷适应蛋白质中具有更高的疏水性。在异亮氨酸的例子中,X射线结构显示,在冷适应酶中,异亮氨酸在亚基界面处的聚集不存在,而在来自超嗜热菌的同源物中则存在紧密堆积的疏水簇。
研究表明,疏水表面残基会由于水分子熵的降低而导致蛋白质结构不稳定,因为这些水分子会在非极性残基周围形成笼状结构。研究发现,在较低温度下,由于释放的水分子迁移率降低,熵增实际上会减少,这表明冷适应酶可以从增加的表面疏水性中获得灵活性和更大的耐受能力。
某些冷适应酶,如胰蛋白酶和β-内酰胺酶(55),表面电荷增加,主要是负电荷。对于某些酶,负电荷相当高,但在活性位点附近也存在一些正电荷。由于水在低温下具有高粘度和高表面张力,破坏氢键网络的能量成本很高。这种能量成本可以通过与具有高介电常数的水分子相互作用的表面带电或极性氨基酸来抵消。这将有助于适当的溶剂化并帮助维持酶的灵活性。通过更好的溶剂相互作用和正电荷,可以在低温下提高灵活性。
-
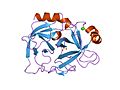 胰蛋白酶的结构
胰蛋白酶的结构

表面斑块中酸性残基的定位可能产生导致蛋白质结构整体不稳定的电荷-电荷排斥。有时会出现酸性残基的电荷排斥可能在连接区产生高水平灵活性的情况。这是某些酶冷适应的主要结构特征。其他研究表明,冷适应涉及溶剂可及和埋藏表面的平均分数减少。这涉及不仅来自古菌家族,也来自细菌家族的其他蛋白质,导致蛋白质表面亲水性增加。通过增加热稳定蛋白质的表面电荷,研究人员能够将其与形成盐桥网络的能力联系起来。这与冷适应酶中与水分子相互作用形成直接对比。已经发现,随着离子相互作用随着温度降低而增强,其数量最小化,从而使冷适应蛋白质在低温下保持灵活性。
Siddiqui KS, Cavicchioli R. 冷适应酶。生物化学年度评论。第75卷:403-33。卷出版日期2006年7月。