
结构生物化学/碳水化合物
碳水化合物是由碳、氢和氧组成的重要大分子。它们是有机化合物,以醛或酮的形式排列,多个羟基从碳链上延伸出来。碳水化合物是生物体内最丰富的有机化合物,占四大生物分子类别之一,包括蛋白质、脂类和核酸。它们是由二氧化碳和水通过光合作用产生的,
nCO2 + nH2O(+ 还原剂和来自光子 [阳光] 的能量)→ ADP + CnH2nOn + nO2
其中 ADP(腺苷二磷酸)是可以通过有氧细胞呼吸合成形成 ATP(腺苷-5'-三磷酸)的产物,ATP 是细胞中使用的化学能形式,作为植物和动物新陈代谢的燃料,
CnH2nOn + nO2 + ADP + 磷酸盐(+ 氧化剂和来自光子 [通过电化学梯度] 的能量)→ ATP + nCO2 + nH2O
碳水化合物在所有生命形式中发挥着各种广泛的作用
碳水化合物的通用经验式为 (CH2O)n。单糖是作为燃料分子以及生命体基本成分的简单糖,是最简单的碳水化合物,作为能量来源是必需的。最常见的是葡萄糖和果糖。
碳水化合物存在于多种异构体形式中。那些原子排列不同的称为结构异构体,例如甘油醛和二羟基丙酮。立体异构体具有相同的原子连接方式,但在空间排列方式上不同,可以进一步分为两种类型:非对映异构体和对映异构体。非对映异构体是非彼此镜像的分子,对映异构体存在为非可叠加的镜像。单糖最多可以拥有三个不同的不对称碳原子这一事实极大地促成了各种各样的异构体形成。
* 作为能量储存、燃料和代谢中间体。
* 形成 RNA 和 DNA(核糖核酸和脱氧核糖核酸)的结构框架。
* 是细菌细胞壁 (肽聚糖或胞壁质)、植物 (纤维素) 和动物 (几丁质) 中的结构元素。
* 它们与许多蛋白质和脂类相连。这种连接的碳水化合物在细胞间通讯以及细胞与细胞环境中其他元素之间的相互作用中很重要。
* 作为能量供应的“食物”(淀粉、糖原、葡聚糖)和脂肪的生产。
对于碳水化合物衍生物,氨基糖是羟基被胺基取代的;脱氧糖是羟基被氢取代的,酸性糖是羧基取代的。
- 在 Hazel M. Holden、Paul D. Cook 和 James B. Thoden 撰写的一篇详细的文章中,揭示了最近对碳水化合物的研究,并提到了对其三维结构的深入发现。碳水化合物占地球生物量的 50%,以其能量储存、植物细胞壁结构和其他功能(如受精和细胞粘附)而闻名。目前的研究正针对微生物和细菌提供的具有化学重要性的新糖类进行。一些微生物糖是二脱氧糖和三脱氧糖,由细菌、真菌和植物制造。一些存在于革兰氏阴性菌或红霉素中。科学家相信这些微生物糖在化学中具有独特的功能,因此,已经进行了对其三维结构的研究。根据最近的研究,微生物产生的糖是由葡萄糖-6-磷酸或果糖-6-磷酸制成的。脱氧糖是通过乙酰化、胺化、差向异构化和甲基化反应的修饰形成的。发现这些糖是从原核生物来源中提取的,它们本身在真核生物系统中具有生物学意义。
- 独特微生物糖的生产始于 NDP-4-酮基-6-脱氧葡萄糖的形成。这种糖是通过生物合成途径形成的,其中 α-D-葡萄糖-1-磷酸通过核苷酰转移酶连接到 NMP 部分,然后是 C-6' 羟基的去除以及 C-4' 羟基部分的氧化。随着 NDP-4-酮基-6-脱氧葡萄糖的形成,会发生重要的酶促反应,例如乙酰化、异构化、脱水、差向异构化等。这些反应负责生产独特的生物合成微生物糖。
- 目前正在研究两种酶,以了解它们在催化 N-乙酰化和 PLP 依赖性胺化中的功能。N-乙酰基转移酶是催化乙酰辅酶 A 到伯胺受体乙酰基转移的酶。这种特定酶的家族成员正在被学者和科学家们严格研究。一个重要的家族成员是 LbetaH,负责通过 O-乙酰基转移酶合成脂质 A。三维研究表明,脂质 A 分子的结构显示出“左手交叉连接及其富含异亮氨酸的六肽基序”。该家族中的另一个重要酶是 PglD,在辅酶 A 复合物中。该酶是一种三聚体,其每个亚基具有两个结构域。发现 PglD 结构的底部是 His 125。这种底部是催化核苷酸连接糖的关键要求。OdtC 是另一种酶,催化 Quip3NAc 制造的最后一步。这种糖存在于一些革兰氏阴性菌中。与 PglD 一样,QdtC 是一种三聚体,但每个亚基都参与了与其他单体的积极相互作用。研究这两种酶的重要性在于,这两种酶具有非常相似的结构,但令人惊讶的是具有不同的生物学功能。详细的结构分析表明,酶可以具有非常相似的结构,但具有完全不同的活性。
- 另一种在酶促反应中起作用的酶,由 NDP-4-酮基-6-脱氧葡萄糖的产生开始,是 PLP 依赖性氨基转移酶和脱水酶。这种酶的名称来源于这样一个事实,即为了成功地将氨基转移到糖上,需要磷酸吡哆醛 5'-磷酸酯 (PLP)。因此,氨基转移酶是 PLP 依赖性的,它被称为 PLP 依赖性氨基转移酶。第一个执行此功能的糖是 ArnB。该糖催化 UDP 连接糖的氨基转移,以产生 UDP-4-氨基-4-脱氧-L-阿拉伯糖。发现 ArnB 执行的胺化是一种“乒乓机制”,其中“乒”是 PMP 的形成,“乓”是它与酮的反应形成“转氨基产物”。在 PLP 反应中,谷氨酸的 -氨基攻击 PLP 的 C4' 使赖氨酸的位置发生偏移。结果,外部亚胺崩溃并形成 -酮戊二酸和 PMP。PMP 的形成为一系列 PMP 与碳水化合物的酮形式之间的反应让路,以通过氨基转移产生产物。
- 在 PLP 依赖性氨基转移酶家族中,PseC 已经过研究,因为研究人员认为其结构在生物学中发挥着重要的作用。他们认为 PseC 可以提供对具有核苷酸连接糖作为底物的氨基转移酶的首次了解。从最近的研究来看,PseC 被发现参与假氨基酸的生物合成,并且还催化碳水化合物 C-4' 位置的氨基转移。另一种重要的酶是 ColD,ColD 可以通过脱水将糖转化为去除 3' 羟基。GDP-过氨基糖合酶也很有趣,因为它使用与 ColD 相同的底物,但它不是执行脱水以去除羟基,而是催化 C4' 位置的胺化。
- 独特微生物糖的生产始于 NDP-4-酮基-6-脱氧葡萄糖的形成。这种糖是通过生物合成途径形成的,其中 α-D-葡萄糖-1-磷酸通过核苷酰转移酶连接到 NMP 部分,然后是 C-6' 羟基的去除以及 C-4' 羟基部分的氧化。随着 NDP-4-酮基-6-脱氧葡萄糖的形成,会发生重要的酶促反应,例如乙酰化、异构化、脱水、差向异构化等。这些反应负责生产独特的生物合成微生物糖。
- 总的来说,许多酶被科学家研究,因为它们在酶促反应中很重要,这些反应产生了细菌和微生物中糖的生物合成。由于酶中的广泛多样性,科学家们力求研究和解释独特微生物糖的形成。
以下是发现 ArnB 的鼠伤寒沙门氏菌的图像。ArnB 是一种已知修饰氨基转移酶结构的第一个糖。 
1. 单糖
2. 二糖
3. 多糖
单糖 (C-H2O)n,n > 3,是碳水化合物的最简单形式。它们可以细分为 醛糖 或 酮糖,如果分子分别含有醛或酮官能团。醛糖在碳链末端具有醛形式的羰基,而酮糖在糖主链的某个位置具有酮基。最简单的酮糖是二羟基丙酮(没有手性中心),而最简单的醛糖是甘油醛,它可以是 L 或 D 对映异构体。单糖可以根据碳原子数进一步分类,例如三碳糖 (n=3)、四碳糖 (n=4)、五碳糖 (n=5)、六碳糖 (n=6)、七碳糖 (n=7) 等。例如,葡萄糖被称为醛己糖(六碳醛糖),而核糖被认为是醛戊糖(五碳醛糖)。糖具有形成环状化合物的能力,通常是五元或六元杂环。此外,糖可以根据其直链形式中是否具有羰基部分来确定是否为还原糖或非还原糖。这意味着环状形式的糖处于半缩醛或半缩酮形式。单糖有助于核酸的形成。它们通常用费歇尔投影式表示。L 和 D 代表单糖上距酮或醛基最远的不对称碳的绝对构型。生物碳水化合物主要是 D 糖。请注意,D 和 L 标记与特定旋转的 D 和 L 标记(右旋和左旋分子)无关。此外,除第一个和最后一个碳原子外,与羟基键合的每个碳原子都是 R 或 S 的立体中心,这可能会为相同的糖类化学式形成许多异构体。在环状形式中,糖可以用霍沃斯投影式描述。
单糖在其环状形式中被归类为 α 或 β 异头物,其中羰基氧的碳原子被称为异头碳原子。由于羰基碳在开链形式中是 sp2 杂化的,当糖环化时,羰基碳形成一个新的手性中心;葡萄糖的 α 异头物在异头碳上连接的羟基相对于杂原子氧处于反位。当羟基与杂原子氧位于同一侧或顺位时,就会出现 β 异头物。两种异头物都处于平衡状态,并且不断地在环状和直链构象之间转换。
异头碳被定义为半缩醛或半缩酮碳,这会导致糖类产生不同的立体异构体形式。异头位置很容易被氧化,并能容易地形成糖苷键。
半缩醛/半缩酮、分子内环化和功能

半缩醛 和 半缩酮 分别是在酸性条件下醛或酮与醇反应时形成的。酸性条件使醛或酮上的羰基碳质子化,然后醇上的羟基亲核进攻羰基碳(亲电体)。在单糖中,它们的物理结构可以以开链或环状形式存在。环状结构在能量上更稳定,并且在葡萄糖、果糖和核糖的情况下更为常见。这些糖有两种可能的环状形成,被称为吡喃糖和呋喃糖形成。吡喃糖环是由醛糖(醛糖)形成的,而呋喃糖环是由 酮糖(酮糖)形成的。吡喃糖是 6 元环,类似于吡喃的结构,而呋喃糖是 5 元环,类似于呋喃。例如,在己糖中,C-5 羟基作为醇(亲核试剂)进攻 C-1 醛羰基碳(亲电试剂)。这导致在 C-1 位置形成分子内半缩醛。为了形成分子内半缩酮,酮己糖的 C-5 羟基进攻 C-2 酮基,形成 5 元环,呋喃糖。

戊糖和己糖可以环化。事实上,许多 5 碳或 6 碳糖,如葡萄糖、核糖和果糖,主要以环状分子形式存在。酮糖和醛糖中的羰基与同一糖分子中的醇基反应。这种分子内反应产生分子内半缩醛或半缩酮。如果形成的环状糖是 5 元环,则它是一种呋喃糖。如果形成的环状糖是 6 元环,则它是一种吡喃糖。只有 5 元环和 6 元环可以形成,因为它们是唯一在能量上有利的环状形成类型。重要的是要注意,当糖经历环状转化时,在此过程中会形成一个异头碳。异头碳是半缩醛或半缩酮碳,在环状糖中可以有两种不同的构型。它可以是 α 或 β,具体取决于 -CH2OH 基团和异头碳上的 -OH 基团的相对位置。α 指两个基团位于相对侧,而 β 指两个基团位于同一侧。
环状糖具有构象异构体。当碳 2 位于平面外时,称为 C-2 内式。它也有椅式和船式构象。由于船式构象存在空间位阻,椅式构象更有利。
总的来说,糖具有许多立体化学性质,例如对映异构体(D 和 L)、非对映异构体、异头物(α 和 β)以及立体化学构型相同但 3D 构象不同的分子(椅式和船式)。

单糖 的结构可以绘制成
1. 费歇尔投影式,它采用直链视角,
上图显示了 D-葡萄糖的费歇尔投影式。

2. 霍沃斯投影式,它将分子显示为环状或平面状,取代基位于环的上方或下方。
上图显示了 α-D-葡萄糖吡喃糖的霍沃斯投影式。
3. 在杂环化合物中,五元环和六元环由于环状结构的空间位阻较小而优于其他环。因此,呋喃糖(戊糖)和 吡喃糖(己糖)环是单糖中主要的环状结构。单糖的环类似于呋喃和吡喃分子,因此得名呋喃糖和吡喃糖。
示例(吡喃是左环,呋喃是右环)

在直链单糖中,醛或酮基与其他羟基反应,其中氧原子与两个碳原子之间形成键,形成半缩醛或半缩酮,从而形成杂环;五原子呋喃糖和六原子吡喃糖形式与其直链结构处于平衡状态。
吡喃糖环可以采取不同的构象。可以是椅式或船式。椅式允许取代基处于两个位置,即轴向或赤道向。轴向的取代基会产生空间位阻,因此较大的取代基通常位于赤道向,因为空间位阻较小。船式构象由于空间位阻而不利。呋喃糖环可以采取信封形式。它有四个几乎共面的原子和一个弯曲的第五个原子。
从费歇尔投影式形成环状结构的基本步骤
1. 以使羟基通过垂直线连接的方式旋转最后一个碳上的取代基。
2. 脊柱右侧的所有取代基,通过水平线与其碳原子连接,将位于环的底部。
3. 脊柱左侧的所有取代基,通过水平线与其碳原子连接,将位于环的顶部。
4. 以顺时针方向将异头碳连接到与最后一个碳相连的羟基。
5. 环状结构通常以霍沃斯投影式(侧面平面视图)绘制为吡喃糖(六元环)或呋喃糖(五元环)。
三碳糖 (n=3,其中 n 指构成糖骨架的碳原子数) 是最小的单糖形式:二羟基丙酮 (酮糖)、甘油醛 (醛糖)
- 单糖示例
- 葡萄糖(最常见的糖)- 葡萄糖是多羟基醛。碳水化合物化学家埃米尔·费歇尔因证明葡萄糖的结构而获得了诺贝尔奖。
- 果糖(使水果变甜)- 果糖是多羟基酮。
- 半乳糖(牛奶中的糖)- 半乳糖是葡萄糖的 C4 差向异构体。与葡萄糖一样,半乳糖在溶于水时发生变旋现象。
- 核糖(RNA 的组成部分)- 阿拉伯糖的 C2 差向异构体,在其呋喃糖形式中最为稳定。每个核糖单元都携带一个杂环碱基,该碱基提供部分信息,以在蛋白质合成中指定特定的氨基酸。
- 单糖示例
***Deoxyribose (component of DNA)-The principal difference between RNA and DNA is the presence of o-2-deoxyribose as the sugar in DNA instead of the o-ribose found.
在 RNA 中。
-
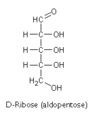 D-核糖
D-核糖 -
 D-果糖
D-果糖 -
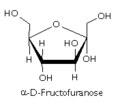 果糖呋喃糖
果糖呋喃糖 -
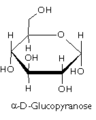 葡萄糖吡喃糖
葡萄糖吡喃糖




二糖,或化学式为 Cn(H2O)n-1,其中 n > 5,即两个通过缩合反应或脱水反应连接的单糖。结果,当一个糖分子的 -OH 与另一个糖分子的 -OH 连接时,会形成糖苷键。糖苷键可以通过水解反应(即添加水分子)来断裂,形成两个单糖。
- 二糖的例子
- 乳糖(牛奶中的糖) - 1 个半乳糖 + 1 个葡萄糖
- 二糖的例子
乳糖由半乳糖通过 β-1,4-糖苷键与葡萄糖连接而成。乳糖酶是人体中将乳糖水解回两种单糖的酶。
- 麦芽糖(麦芽糖) - 1 个葡萄糖 + 1 个葡萄糖
麦芽糖由两个通过 α-1,4-糖苷键连接的葡萄糖分子构成。麦芽糖酶是水解麦芽糖的酶。
- 纤维二糖 - 1 个葡萄糖 + 1 个葡萄糖
纤维二糖也是由两个葡萄糖分子构成。与麦芽糖不同,它通过 β-1,4-糖苷键连接,由于赤道键的存在,它是一种不可消化的糖。纤维二糖是纤维素的二糖,并且是一种还原糖,因为它具有半缩醛。
- 蔗糖(食糖) - 1 个葡萄糖 + 1 个果糖
蔗糖由葡萄糖和果糖构成。它们在 α 葡萄糖和 β 果糖之间形成糖苷键。由于它们在端基碳原子上连接,因此蔗糖不是还原糖,因为葡萄糖或果糖都不能转化为醛或酮。
一般来说,糖苷键是指两个单糖之间形成二糖的共价键。它是在酸性条件下,通过缩合反应,在糖的端基碳原子的半缩醛基团与有机化合物的羟基之间形成的。缩合反应导致形成一个额外的水分子。这导致在端基碳原子上形成一个缩醛基团,从而稳定了该键。糖苷键非常稳定,但可以在强酸性水溶液中通过化学方法断裂。糖苷键有两种类型:α 糖苷键和 β 糖苷键。具有糖苷键的物质被称为糖苷。
对于糖类来说,如果它们具有端基碳原子,则可以形成糖苷键以形成环状结构,而不是线性形式。该化学键被认为是 O-糖苷键(II 型连接),其中端基碳原子通过氧原子与其他基团连接。O-糖苷键在丝氨酸或苏氨酸的羟基侧链酰胺与糖之间形成共价连接。N-糖苷键,I 型连接,是一种碳水化合物-蛋白质共价连接类型,存在于天冬酰胺的侧链酰胺与糖之间。N-糖苷键涉及糖的端基碳原子与胺基的氮原子之间的连接。
在形成糖苷键的过程中,糖类的半缩醛基团(含有端基碳原子)在酸的存在下形成糖苷键。费歇尔糖苷化是该缩合反应的结果,该反应会从之前的反应中释放出一个水分子。
尽管糖苷键在大多数条件下相对稳定,但在许多酸性水溶液中很容易断裂。还有一些被称为糖苷水解酶的酶也可以断裂这些键,但它们只能断裂 α 或 β 型构象,不能断裂两种构象。下图展示了生成糖苷键的反应示例。http://en.wikipedia.org/wiki/File:Ethyl-glucoside.png
键的立体化学基于最远离 C1 的立体异构中心的位点。糖苷键的 α 型构象形成轴向构象,而 β 型构象形成赤道构象。
寡糖是多糖,其中糖分子数量较少;二糖与更简单的多糖连接,其中 n=3 或 4 个简单糖单元。由于几乎所有真核细胞的分泌和膜相关蛋白都发生糖基化,因此寡糖通过 N-糖苷键或 O-糖苷键共价连接到蛋白质。
寡糖主要有两种类型
- N-连接的寡糖:在 N-连接的寡糖中,GlcNAc 以可变的 β 连接方式连接到 Asn 残基中 Asn-X-Thr 或 Asn-X-Ser 序列的酰胺 N,其中 X 是除可能为 Pro 或 Asp 以外的任何氨基酸。N-连接的糖蛋白中的寡糖具有巨大的多样性。在某些糖蛋白中,加工过程很简短,留下“高甘露糖”寡糖。在其他一些糖蛋白中,广泛的加工会产生含有几种糖残基的大型寡糖。N 连接的寡糖具有一个共同的五糖核心,该核心含有 3 个甘露糖和 2 个 N 乙酰葡糖胺残基。
含有 N-连接的寡糖的蛋白质通常会发生糖基化,然后按如下方式进行加工,这在很大程度上得到了阐明
1. 一个含有 9 个甘露糖残基、3 个葡萄糖残基和 2 个 GlcNAc 残基的寡糖连接到正在由与内质网相关的核糖体合成的生长中的多肽链的 Asn 上。
2. 在该过程中,一些糖被去除,该过程从内质网腔开始,并在高尔基体继续进行。
3. 通过高尔基体中特定糖基转移酶的作用,添加了额外的单糖残基,包括 GlcNAc、半乳糖、岩藻糖和唾液酸。
- O-连接的寡糖:最常见的 O-糖苷连接涉及二糖核心 β-半乳糖基-(1->3)-α-N-乙酰半乳糖胺,该核心连接到丝氨酸或苏氨酸的 OH 基团。O-连接的寡糖在高尔基体中通过将单糖单元连续添加到已完成的多肽链中来合成。合成从将 GalNAc 转移到多肽上的丝氨酸和苏氨酸残基开始。N-连接的寡糖被转移到特定氨基酸序列中的 Asn,但 O-糖基化的丝氨酸和苏氨酸残基不是任何常见序列的成员。相反,糖基化位点的定位仅由多肽的二级或三级结构决定。O-糖基化继续进行,通过相应的糖基转移酶逐步添加糖。
内质网 (3) = N-连接的高尔基体复合体 (8)= O-连接和 N-连接

- 寡糖的测序:为了确定寡糖的结构及其连接点,Berg 等人[1]指出,第一步是将寡糖从蛋白质上分离,然后将寡糖分离并进行分析。Berg 等人指出,质谱技术可以提供寡糖片段的质量。Berg 等人给出的一个例子是,通过使用诸如肽 N-糖苷酶 F 之类的酶从蛋白质中释放 N-连接的寡糖。N-糖苷酶 F 切断将寡糖连接到蛋白质的 N-糖苷键。


多糖(也称为聚糖),化学式写为 Cn(H2O)n-1,其中 200 < n < 2500,或 (C6H10O5)n,40<n<3000,是单糖的聚合物或多个单元。它们通过糖苷键构建,由于其结构和溶解度,它们是理想的储存产物。寡糖是糖分子数量较少的多糖;二糖与更简单的多糖连接,其中 n=3 或 4 个简单糖单元。寡糖通过 N-糖苷键或 O-糖苷键共价连接到蛋白质。多糖可以根据单个单糖的组成进一步分类为同多糖和异多糖。此外,支链和无支链多糖是通过单糖的不同碳原子上的糖苷键形成的。
同多糖由一种类型的单糖组成。它们可以是支链的或无支链的。异多糖可以由直链或支链中的两种或多种类型的单糖组成。同多糖的例子:1)一些是能量储存(淀粉、糖原和葡聚糖)。2)其他具有结构作用(纤维素、几丁质)。
多糖的例子
- 纤维素
纤维素 [(C6H10O5)n] 是一种同多糖,在植物细胞壁中起着重要的结构作用。例如,棉花纤维和树木和灌木的木材主要由纤维素组成。纤维素中的葡萄糖残基以线性方式通过 β-1,4-糖苷键连接。分子间和分子内氢键使纤维素的疏水部分堆叠,导致纤维素在正常水溶液中完全不溶。
- 淀粉
淀粉是葡萄糖的聚合物,由通过 α-糖苷键连接的糖吡喃糖单元构成。淀粉是一种同多糖,是植物中主要的能量储存来源,也是包括人类在内的大多数动物的营养来源。直链淀粉和支链淀粉是淀粉碳水化合物的例子。直链淀粉是通过 α-糖苷键连接的数千个葡萄糖残基的线性链,而支链淀粉是多糖的支链。直链淀粉的线性结合是 α-1,4-糖苷键。支链淀粉的支链是 α-1,6-连接。支链淀粉中的分支比糖原少。这些分支大约每 30 个 α-1,4-连接出现一次。葡萄糖的聚合物不溶于水。
- 糖原
糖原,也称为“动物淀粉”,是动物细胞储存葡萄糖作为能量来源的方式。它的结构类似于支链淀粉,但分支更多。糖原由葡萄糖吡喃糖单元链组成,这些单元通过 α-1,4-糖苷键连接,聚合物的分支是由于 α-1,6-糖苷键。每十个 1,4-连接就会出现一次分支。糖原主要在肝脏和肌肉细胞中产生。
- 几丁质
几丁质是一种同多糖,由重复的葡萄糖衍生物 N-乙酰葡萄糖胺单元组成。这些单元通过 β-1,4-连接线性连接。几丁质是一种非常重要的**结构性**成分,构成真菌的细胞壁、节肢动物和昆虫的外骨骼以及其他此类成分。它的结构和连接与纤维素相似,除了葡萄糖 2' 碳上的羟基被乙酰胺基取代。
结构差异
淀粉和糖原中通过 α-1,4-糖苷键连接赋予它们与纤维素(通过 β-1,4-糖苷键连接)完全不同的结构。α-1,4-糖苷键导致空心螺旋结构,更适合能量储存,而 β-1-4-糖苷键允许纤维素形成线性链,这些线性链通过与相邻链的氢键稳定,形成抗拉纤维。
-
 纤维素
纤维素 -
 淀粉
淀粉 -
 糖原
糖原 -
 几丁质
几丁质



{kind=link}
糖的类型
[edit | edit source]糖也可以被归类为**还原糖**或**非还原糖**。

还原糖
[edit | edit source]
还原糖可以获得其开链形式。还原糖基本上是具有醛基(在其开链形式中)或**半缩醛**基(在其环状形式中)的糖,该基位于准备氧化的异头碳上。换句话说,还原糖允许链形成和延伸。可以使用斐林试剂(一种含 Cu2+离子的溶液,可以很容易地氧化游离醛或酮)来测试还原糖的存在。可以使用贝内迪克特试剂进行类似的测试,该试剂也包含关键的 Cu2+ 离子。还原糖的一个特点是 β-糖苷键,其中 β 异头碳参与连接。还原糖的例子包括葡萄糖、麦芽糖和乳糖。在碱性条件下,还原糖可以形成醛基或酮基,因为羧基被还原为醛基或酮基的羰基,并且当异头碳不与半缩醛或半缩酮羟基键合时发生。
仅提到醛糖可以形成还原糖。酮糖怎么样?从机理上讲,当酮糖异构化为其环状形式时,会形成缩醛。所以,酮糖不会形成还原糖。然而,酮糖可以互变异构为醛糖,然后在环闭合时可以形成半缩醛。[2]
非还原糖
[edit | edit source]
非还原糖无法获得游离的醛或酮,例如糖苷。这种类型的糖基本上是在异头碳上的**缩醛**(在其环状形式中)。由于异头碳固定在糖苷键中,因此糖链无法形成或延长。非还原糖也可以通过 α-糖苷键来识别,其中 α 异头碳参与连接。非还原糖的一个例子是蔗糖。
糖蛋白
[edit | edit source]- 结构
糖蛋白,即碳水化合物和蛋白质混合物的有机化合物,通过共价键结合,是在**糖基化**过程中形成的。碳水化合物基团可以共价键合到蛋白质上形成糖蛋白,它是细胞膜的重要组成部分,参与诸如细胞粘附和受精过程中精子与卵子的结合等过程。在糖蛋白中,碳水化合物的重量百分比远小于蛋白多糖中发现的碳水化合物的重量。糖蛋白的糖基团有助于蛋白质折叠并增加蛋白质稳定性。糖蛋白中的糖以两种方式连接:通过**N-连接糖苷键**连接到天冬酰胺侧链的酰胺氮原子上,或通过**O-连接糖苷键**连接到丝氨酸或苏氨酸侧链的氧原子上。碳水化合物和蛋白质之间的键合是通过与天冬酰胺、羟赖氨酸、丝氨酸或苏氨酸的四个氨基酸之一的键合形成的。因此,可以预测蛋白质糖基化位点,因为天冬酰胺只有在以下形式中才能被糖基化:Asn-X-Ser、Asn-X-Thr 或 Asn-X-Cys 序列,其中 X 可以是任何氨基酸,但脯氨酸除外。但并非所有潜在的位点都被糖基化。糖基化位点具有其他因素,例如蛋白质表达的细胞类型和蛋白质结构。在 N-连接寡糖中,它具有一个常见的五糖核心,包含三个甘露糖和两个 N-乙酰葡萄糖胺残基。为了形成各种各样的寡糖模式,额外的糖被连接到这个核心。糖蛋白的整合膜蛋白对于一个细胞与另一个细胞之间的相互作用很重要。
- 糖基化
蛋白质胞外部分的糖基化发生在内质网和高尔基体中。内质网和高尔基体是参与蛋白质转运/包装的主要作用的细胞器。高尔基体是细胞的主要分类中心。高尔基体改变并添加糖蛋白的碳水化合物单元,因为它细胞的主要包装/分类中心。蛋白质从高尔基体到溶酶体、分泌颗粒或质膜。粗面内质网的胞质面上核糖体合成被带入内质网腔的蛋白质。N-连接糖基化从内质网开始,并在高尔基体中继续。它始于添加由 14 个糖分子链组成的寡糖前体。然而,O-连接糖基化位点仅在高尔基体中。此外,与 N-连接糖基化不同,O-连接糖基化一次添加一个糖分子,每个糖分子由不同的糖基转移酶添加。一个例子包括在高尔基体的顺式区域,通过 N-乙酰半乳糖胺转移酶添加 N-乙酰半乳糖胺 (GalNAc)。(应该注意的是,高尔基体被分成三个区域,即顺式、反式和中间式)。在高尔基体的反式区域,通过高尔基体该区域特有的半乳糖胺转移酶,将半乳糖残基连接到 N-乙酰半乳糖胺上。要连接到蛋白质中天冬酰胺残基酰胺侧链上的寡糖前体首先连接到多萜醇磷酸。多萜醇磷酸是内质网腔中发现的一种脂类分子,由大约二十个异戊二烯单元组成。多萜醇磷酸的末端磷酸基团是寡糖连接的位置。在寡糖-蛋白质转移酶的帮助下,寡糖从多萜醇磷酸转移到天冬酰胺分子。然后,来自内质网腔和内质网膜的蛋白质被转移到高尔基体,在那里糖蛋白的碳水化合物部分被改变。由于高尔基体有三个区域,每个区域都有自己的一组酶,因此对前体寡糖的修饰允许形成一系列的寡糖结构。在高尔基体之后,蛋白质根据嵌入氨基酸序列和三维结构中的信号,继续到达溶酶体、分泌颗粒或质膜。
- 例子
促红细胞生成素 (EPO) 是一种糖蛋白激素,可刺激红细胞的生成。三个 Asn 残基和一个 Ser 残基的存在允许寡糖在三个 N-连接糖基化位点和一个 O-连接糖基化位点连接蛋白质。它由肾脏分泌。成熟的 EPO 包含约 40% 的碳水化合物,通过糖基化,它将增强其蛋白质的稳定性。当 EPO 未经糖基化时,其蛋白质会迅速从血液中清除,因此仅包含 10% 的碳水化合物。

透明带是一种糖蛋白膜,出现在质膜周围的多层原始卵母细胞中。透明带结构必须引发顶体反应,以便与精子结合。因此,科学家们发现了四种透明带,它们负责小鼠精子与顶体反应的结合。最重要的透明带糖蛋白是 ZP3,因为 ZP3 负责精子结合。精子蛋白与卵母细胞的质膜粘附。此外,ZP3 还参与顶体反应;这导致精子释放顶体囊泡。ZP2 负责介导随后精子的结合。因此,由于 ZP1,ZP2 和 ZP3 交联。ZP4 是人类编码基因的蛋白质。对于人类来说,受精后五天,囊胚进行透明带孵化。另一方面,当透明带分解和退化时,透明带被滋养层细胞层取代。总的来说,透明带对卵子死亡和受精的开始具有重要意义。
- 寡糖序列
可以使用酶分析和质谱法对寡糖进行测序。很难知道糖的结构,因此需要从糖蛋白中去除糖。您将使用酶和质谱法来找出这些附着糖的顺序。
- 糖基化错误
碳水化合物与蛋白质的结合对于加工、稳定性和靶向这些蛋白质至关重要。蛋白质的糖基化不当会导致称为糖基化先天性疾病的遗传性人类疾病。一个例子是 I-细胞疾病。I-细胞疾病是一种溶酶体贮积病。碳水化合物标记用于引导降解酶。患有 I-细胞疾病的人的溶酶体中含有大量未消化的糖胺聚糖。这些包涵体存在是因为 I-细胞患者的溶酶体缺乏降解它们的酶。然而,这些酶在身体的其他部位大量存在,因此表明 I-细胞患者的酶传递错误。
碳水化合物结合蛋白

研究表明,碳水化合物-蛋白质复合物在细胞-细胞识别过程中发挥作用,以及细胞与邻近细胞和细胞外基质的粘附。细胞表面展示的各种碳水化合物结构非常适合作为细胞与其环境之间相互作用的位点。当碳水化合物基团通过共价键连接到蛋白质时,就会形成糖蛋白。这些糖苷键分别将碳水化合物连接到天冬酰胺和丝氨酸或苏氨酸的氨基和羟基侧链。N-连接是碳水化合物与天冬酰胺侧链中氮之间的键,O-连接是碳水化合物与丝氨酸或苏氨酸氧之间的键。只有当天冬酰胺残基是 Asn-X-Ser 或 Asn-X-Thr 序列的一部分时,它才能接受寡糖,其中 X 可以是任何氨基酸,除了脯氨酸。因此,可以在蛋白质的一级结构中检测到潜在的糖基化位点。然而,并非所有潜在位点都发生糖基化。糖基化位点取决于该区域内的蛋白质结构以及蛋白质表达的细胞类型。所有 N-连接的寡糖都有一个共同的五糖核心,该核心由三个甘露糖和两个 N-乙酰氨基葡萄糖残基组成。
糖蛋白在医学领域发挥着多种作用。修饰的碳水化合物能够干扰碳水化合物与蛋白质之间的相互作用。这导致细胞-细胞识别和粘附的抑制,这是导致癌细胞生长的主要因素。因此,这些碳水化合物结合蛋白的配体可能会演变成新的癌症治疗形式。人们一直在研究基于蛋白质血清的癌症诊断方法。类似地,来自肾脏的糖蛋白激素红细胞生成素改善了贫血的治疗,通过刺激红细胞的增加。此外,膜蛋白的正确糖基化通过稳定钾离子通道来防止人类组织和肌肉萎缩的降解。
凝集素
植物和动物中特异性的碳水化合物结合蛋白是凝集素,它们是结合碳水化合物结构并促进细胞-细胞相互作用的伙伴。典型的凝集素包含 2 个或更多个碳水化合物结合位点,并表现出蛋白质与碳水化合物之间的弱相互作用。这些多个弱相互作用累加在一起形成一个强烈的连接。凝集素的连接就像魔术贴一样,弱相互作用但强烈的复合物!凝集素存在于动物、植物和微生物中。凝集素在细胞识别中也发挥着重要作用,因为修饰的碳水化合物和寡糖能够干扰碳水化合物-蛋白质相互作用,从而抑制细胞-细胞识别和粘附过程。凝集素具有高度的专一性:它们可以识别寡糖中特定连接到其他糖的单个单糖(这种特性也使凝集素成为分离糖蛋白和寡糖的有用实验室工具)。一个细胞表面的碳水化合物与另一个细胞表面凝集素的结合位点结合。
- 碳水化合物在细胞通讯中的重要性
由于可能的各种组成和结构,碳水化合物已被证明能够包含大量信息。这些多样化的化合物是由单糖含有许多可用于连接的 OH 基团这一事实造成的。这进一步允许广泛的分支。此外,连接到端基碳的取代基可以是 α 或 β 型。这些各种碳水化合物在细胞表面的存在允许有效的细胞间通讯。
- C 型凝集素

C 型凝集素是那些需要钙离子的凝集素。钙离子通过与碳水化合物上的 OH 基团相互作用来帮助结合蛋白质和碳水化合物。钙还可以形成碳水化合物与凝集素中的谷氨酸之间的连接。通过凝集素侧链与碳水化合物 OH 基团之间形成的氢键进一步增强结合。碳水化合物识别和结合是通过一个由 120 个氨基酸组成的同源域实现的。这些氨基酸决定了碳水化合物结合的专一性。
- 凝集素的例子
胚胎通过 L-选择素附着在子宫内膜上。这激活了一个信号以允许植入。大肠杆菌能够通过识别肠道中碳水化合物的凝集素驻留在胃肠道中。
麦芽凝集素
麦芽凝集素是一种存在于细胞内质网中的碳水化合物结合蛋白。它在 N-糖基化的早期阶段起着重要的作用。
选择素
选择素是形成单链的跨膜糖蛋白。它们介导白细胞(循环白细胞)与内皮细胞(血管内壁细胞)表面的附着。选择素是 C 型凝集素的成员。选择素识别并结合细胞表面糖蛋白上的特定寡糖。两种细胞类型之间的选择素-寡糖相互作用使内皮细胞能够“捕获”循环白细胞,然后白细胞沿着内皮细胞爬行,以消除感染或帮助修复受损组织。选择素将特定的免疫细胞与损伤和炎症部位结合。选择素有各种类型。例如,L 型选择素指的是淋巴结血管,E 型选择素指的是内皮,P 型指的是血小板。
病毒
粘附到细胞表面碳水化合物使一些病毒能够进入细胞。病毒可以结合糖蛋白上的特定残基,一旦进入细胞,就会释放蛋白质,这些蛋白质会破坏糖蛋白中的某些键,使病毒可以自由进入。流感病毒结合唾液酸残基,唾液酸残基存在于细胞表面糖蛋白上。结合硅酸残基的病毒蛋白被称为血凝素。病毒穿透细胞膜后,另一种病毒蛋白神经氨酸酶(唾液酸酶)会切割唾液酸残基的糖苷键,使病毒可以自由感染细胞。这种酶的抑制剂,如达菲和瑞乐沙,是重要的抗流感药物。

碳水化合物的多样性
由于以下原因,碳水化合物非常多样化
1. 不同的单糖可以通过几个 OH 基团中的任何一个连接在一起。
2. C-1 连接可以是 α 型或 β 型,并且可以进行广泛的分支。
与寡肽相比,可以用四种糖形成许多不同的寡糖。
由于碳水化合物化学性质导致的多样性
化学结构。
单糖由于其大量的立体异构中心,是天然的复杂构建块。C6H12O6(己糖)有 16 个立体异构体。其中两个是 D- 和 L-半乳糖的对映异构体(D-Gal 和 L-Gal)。在人类中,只有 D-Gal 存在,而在植物中,D 型和 L 型都可以找到。游离的单糖是多羟基醛或多羟基酮,可以以开放形式存在,也可以发生分子内反应产生环状半缩醛。进攻的 OH 决定了环的大小,通常是六元环(吡喃糖环)或,较不常见的是,五元环(呋喃糖环)。由于环化,形成一个新的手性碳,产生两种可能的异构体(α 和 β),它们与开放形式处于平衡状态。这些半缩醛可以反应形成缩醛,固定端基碳的立体化学,产生各种碳水化合物衍生物。例如,D-Gal 在某些微生物中以吡喃糖和呋喃糖形式存在,即使在同一糖缀合物中也是如此。
化学连接性:糖苷键和分支。
由于单糖之间可用于形成分子间缩醛的羟基不同,因此一个单糖的异头碳与另一个单糖的任何羟基之间都可能形成许多糖苷键。这种反应决定了键的类型(α-键或β-键),这对于酶识别至关重要。不同分支点的可用性是影响碳水化合物紧凑性、柔性和物理和生化性质的关键因素。一个明显的例子是淀粉和糖原之间的区别——淀粉由直链淀粉组成,直链淀粉是α(1,4)-葡萄糖残基的线性同聚物,在植物中以半结晶颗粒形式沉积;而糖原也是α(1,4)-葡萄糖残基的同聚物,但每十个单位中存在一个α(1,6)分支点,因此它以非晶态多糖的形式存在于动物细胞中。
非碳水化合物修饰
糖链结构可以通过多种取代进行修饰,包括硫酸化、乙酰化或磷酸化。
- ↑ Berg J. M., Tymoczko J L., Stryer L. “生物化学” 第六版, (c) 2007, 2002 by W. H. Freeman and Company, New York
- ↑ http://www.chem.ucalgary.ca/courses/350/Carey5th/Ch25/ch25-2-5.html
1. Berg, Jeremy M. 2007. 生物化学. 第六版. 纽约: W.H. Freeman. 303-325.
2. "选择素家族的碳水化合物结合蛋白:结构和碳水化合物配体对细胞粘附的重要性". Richard D. Cummings, David F. Smith. 生物论文集. 第 14 卷,第 12 期,第 849-856 页,1992 年,Wiley Periodicals, Inc.
3. "碳水化合物结合蛋白在治疗各种疾病方面正变得非常有用", 2004 年 4 月,News-Medical.Net [2]
4. Nangia-Makker, P.; Conklin, J.; Hogan, V. 以及 Raz, A. "癌症中的碳水化合物结合蛋白及其配体作为治疗剂", 分子医学趋势. 第 8 卷,第 4 期,2002 年 4 月 1 日,第 187-192 页
5. Richards, M.R. & Lowary, T.L. 含有半乳糖呋喃糖的聚糖的化学和生物学. ChemBioChem 10, 1920–1938 (2009).
6. Wade, L. G. "23 碳水化合物和核酸." 有机化学. 第 7 版. 上鞍河,新泽西州:Pearson Education,2009。1097-141. 印刷版。
7. Holden, Hazel M., Paul D. Cook, 以及 James B. Thoden. “不寻常微生物糖的生物合成酶”. 结构生物学当前观点. (2010): 543-550. ScienceDirect.