
结构生物化学/进化基础
进化与遗传学研究密切相关,观察生物体从一代到下一代的遗传和基因的变化。这在真核生物的进化中得到了最好的体现,导致了动物和人类等更复杂的生物体的出现。
进化伴随着基因中微小的变化,称为突变。生物体具有生存的本能能力。因此,突变的发生是为了满足自然选择,只有有利于生存的性状才能遗传给后代。进化的另一种机制被称为遗传漂变。这个过程独立发生,导致群体性状的随机变化。这是导致新物种出现的主要机制。
由于生物化学研究基因中发生的突变以及某个基因、细胞或生物分子与另一个基因、细胞或生物分子之间的关系,生物化学研究与遗传学合作,构建进化树,使科学家能够看到不同生物体之间的关系以及它们之间的亲缘关系。
进化是指地球上的生命从其最早形式到今天所表现出的巨大多样性的转变过程。达尔文探讨了生物体的多样性,它们的起源和关系,它们的异同,它们的地理分布以及它们对周围环境的适应性。
大约 35 亿年前,生命起源于地球,第一个生命形式很可能是一个包含自我复制 大分子 分子的膜封闭细胞。推测认为,第一个细胞的成分可能是在海底热液喷口或其他高温区域产生的,那里存在着二氧化碳和氨等大气分子。一个著名的实验表明,生物分子可以从无机成分中形成,这就是米勒-尤里实验。这两位科学家在一个烧瓶中模拟了早期地球的环境,发现 大分子 的单体基础被形成了。
前生物理论试图解释生命的起源,更具体地说,是从无机化学物质中创造出复杂的有机化学结构。最早的化学解释源于奥巴林的著作《生命的起源》。奥巴林的理论因将起点设定为一个可通过实验验证的前生物理论而引人注目。大多数基于生物化学的前生物理论都源于奥巴林原始理论的修改或修订。米勒-尤里实验模拟了奥巴林假设设定的条件,并提供了支持前生物理论的可验证数据。
奥巴林理论[1]关于生命从无机物质演化的理论被称为前生物理论。根据奥巴林的说法,早期大气与今天的大气截然不同。大气是高度还原性的,由甲烷、氨、氢气和水组成。在地球大气层保护能力较弱的情况下,未过滤的太阳辐射的作用会使还原性化学物质发生反应。这些反应会使化学物质相互攻击,形成有机分子,这些分子会由于各种分子相互作用而聚集在一起,形成“原始汤”。在“原始汤”中,有机分子会形成团聚体,即通过疏水力吸引在一起的各种化学化合物;这些团聚体将允许形成半封闭系统,其中可以形成基本的生物代谢。化学反应的复杂性和规模会不断增加,最终形成 氨基酸。这些反应被假设为现代蛋白质结构的起源。然而,奥巴林的理论存在一些问题。奥巴林指出,大气是高度还原性的,因为氧化攻击会破坏更复杂的氨基酸。然而,如果没有氧化气体,臭氧层就无法阻止过量的太阳辐射,而这些辐射最终会将氮气分解成各自的成分。在某种程度上,团聚体提供的包容性是足够的,但它并不是对前生物理论的完整解释或解决方案。
- 大气成分
早期地球的大气与今天的大气有很大不同。假设认为,地球的大气已经进化了许多次,才形成了现在的大气。今天的大气主要由碳、氮、氧和氢组成。关于早期大气的最流行理论是,它几乎不含氧气,主要由氨、甲烷、氢气和水组成。在闪电、火山爆发和紫外线辐射等自然力量的帮助下,大气得以进化。早期大气中被认为含有大量的氢气,这将解释大气是如何通过氧气还原而进化的。米勒-尤里实验能够检验这一假设。该实验仅使用被认为存在于大气中的元素,并用电火花来模拟闪电,证明了地球的大气有可能进化成今天的样子。米勒-尤里实验为整个大气进化过程中形成生物体铺平了道路。该实验产生了气态和水相。气相产生了二氧化碳和一氧化碳,而水相包含醛类、氨基酸和羟基酸等各种化合物。这表明非生物合成生物分子的可能性是存在的。随着更详细的实验的进行,数据表明,在这样的环境下,可以产生多肽和类似 RNA 的分子。也有证据表明,生命可能只在早期发展起来。像涉及氨基酸的那些化学反应需要氧气含量非常低。早期大气也解释了厌氧生命形式的存在。许多实验得出的结论是,大气中的氧气含量随着时间的推移而增加。
2006 年,提出了一种有机化合物的替代来源:外太空。星尘号太空任务的研究带回了从彗星尾巴收集的微小尘埃颗粒,这些颗粒包含各种有机化合物。
- 原始海洋/汤
原始海洋是指地球上所有被认为是多细胞生物体发展开端的海洋的集合。在早期地球演化的过程中,许多有机分子存在,但无法自我复制。然而,由于当时的多细胞生物体尚未出现,有机分子能够以某种方式为化学进化提供适宜的环境。复杂的聚合物也能够在这样的环境中形成,但这种形成并非一个简单的过程。原始汤也指的是米勒-尤里实验,因为它涉及创造一个被认为存在于早期时代的海洋。
1953 年,斯坦利·米勒和哈罗德·尤里(当时在芝加哥大学)在尤里的实验室中设计了一个实验,模拟了奥巴林假设认为存在于早期地球的条件。该实验由一个封闭的系统组成,该系统既有液相,也有“大气”相(见下图)。在“大气”球中,有 NH3、CH4 和 H2 气体(分别为氨、甲烷和氢气),据认为,由于火山活动频繁,它们在早期地球的大气中很常见。液相包含被煮沸以产生水蒸气的水。这种蒸汽会流入气体容器,并暴露在电极产生的电火花中。这些火花模拟了早期地球上常见的频繁闪电。该球中的气体通过冷凝器不断循环,模拟降雨。化合物将从“大气”阶段中移除,并收集回液相中,作为原始海洋的代表。实验条件保持了很长一段时间,有时甚至长达一周或更长时间。定期从“海洋”阶段采集样品进行分析。
对从系统中采集的样本进行分析发现存在有机化合物。发现的化合物包括HCN、醛类、羟基酸、氨基酸和简单核苷酸;其中包括丙氨酸和谷氨酸。还发现气相中含有高浓度的CO和CO2。这些化合物的存在使米勒和尤里得出结论:在假设存在于早期地球的条件下,强还原环境中可以无机地形成早期生命和原生命所必需的有机化合物。
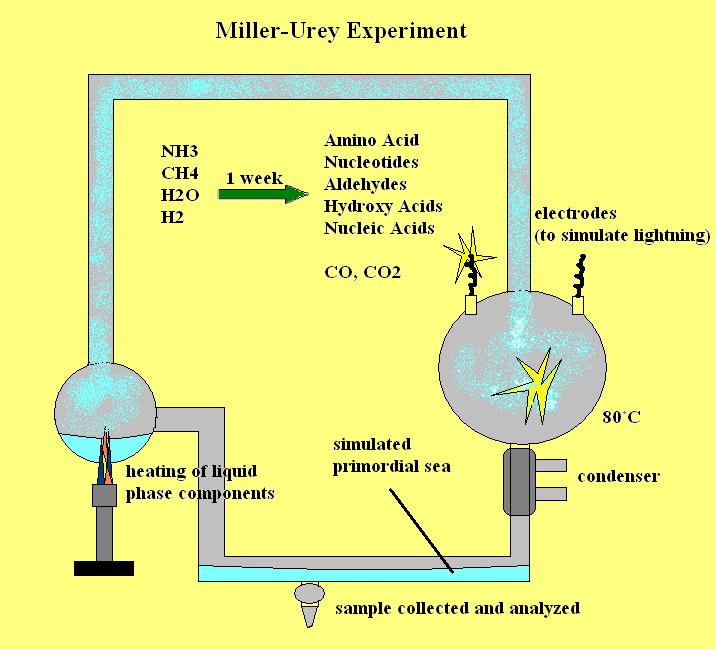

A schematic view of the famous Miller-Urey (AKA Urey-Miller) Experiment.
主要文章: RNA世界假说.
biochemical theory is a mutual dependence between nucleic acids that encode genetic information and the enzymes that catalyze their replication. However, RNA or Related precursors may have been the first genes and catalysts, preceding both DNA and proteins. RNA developed over time from a primordial soup of the early Earth. Nucleotides were one of the components of this atmosphere, and eventually sequenced spontaneously into an early form of RNA. RNA was discovered to have catalytic properties with the possibility of storing genetic information and self replication. Mutations over time could have enhanced these self replicating properties of RNA and perpetuate them further. Therefore, more RNA molecules formed and evolved exponentially to create more efficient peptide sequences and ways of replication. Soon, new variants of self replicating RNA developed, again through mutations, with the ability to catalyze the condensation of amino acids into peptides. Eventually, one of these peptides would form and assist the self replicating ability of RNA generating increasingly efficient self replicating systems. The fact that ribosomes, RNA molecules, not proteins, catalyze the formation of peptide bonds is consistent with this theory. DNA has the complementary sequence to RNA, and so it is thought to have evolved from RNA and took over its function of conserving the genetic information. DNA took over RNA's role as the storage of genetic information because its structure is more stable than that of RNA. These characteristics lead to the hypothesis that RNA led to DNA and protein formation.
人们认为,RNA而不是DNA是第一种遗传物质。这些原始的RNA,尽管短小简单,却能够自我复制,催化这一过程,以及剪接自身的内含子。随着RNA的大小和复杂性的增加,它获得了自我折叠的能力。由于RNA因不断增长的复杂性而变得更低效,突变率也随之增加。由于RNA在可遗传信息的领域占主导地位,这个先于现在的时代被称为“RNA世界”。

RNA表现出极强的耐久性,这对遗传信息的完整性至关重要。然而,随着DNA的出现,RNA的稳定性相形见绌。RNA的核糖环包含两个醇官能团,这使得RNA更容易受到碱催化水解的影响,而DNA只有一个,这反过来又提高了它的稳定性。RNA逆转录成DNA是使用一种特定的酶完成的,这种酶与HIV病毒用来将其病毒RNA逆转录成DNA并将其整合到宿主遗传成分中的酶非常相似。
利用生物化学知识,遗传学家还发现DNA可以特化,不同的功能占据基因的不同部分。有些包含传递信息的代码,而另一些可能包含关于繁殖的信息等等。特化不仅使细胞更加高效,而且也更加可预测。在诸如医学之类的遗传学应用中,研究人员已经能够识别出导致相应脆弱性的基因,范围从生理到心理。然后,基因组可以作为历史记录,跟踪物种随着时间的推移而发生的改变。
虽然DNA成为蓝图,但蛋白质承担着细胞活动调节器和指挥者的角色。当两种蛋白质来自具有相似序列的两个不同基因时,它们被称为同源物。当这些蛋白质出现在同一种类中时,同源物可以进一步细分为两个亚基,即旁系同源物,当它们出现在不同物种中时,被称为直系同源物。通过研究这些现象,科学家可以确定不同物种的进化路径,因为这些发现对于构建系统发育非常有价值。
- 复杂的内部结构,存在高度的化学复杂性和微观组织。数千种分子使细胞能够维持非常复杂的内部结构,其中包括长聚合物,每个聚合物都具有亚基的特征序列、其独特的3D结构以及特定选择结合位点。
- 系统能够通过从环境中提取、转化和利用能量来维持平衡。这使得生物体能够进行机械、化学和渗透性工作。
- 繁殖,具有明确的生物体成分功能,生物体相互作用生物体之间的相互作用使成分发生改变,从而导致协调和补偿,从而适应环境的必要变化。
- 用于感知和响应生物体环境变化的机制不断调整内部化学物质,以适应其局部环境的变化,以获得更好的生存能力。
- 精确的自我复制和自我组装能力构建每个生物体独有的每个细胞的完整遗传物质。
- 随时间推移的进化能力这种生物体的基本统一性反映在基因序列和蛋白质结构的相似性上,这体现在分子水平上。
虽然米勒-尤里实验确实提供了关于诸如氨基酸和核酸之类的有机单体的可能起源,但在实验中收集的样本中没有发现复杂聚合物的证据。
一些科学家推测,某些聚合物(即多肽和小蛋白质)的形成可能是由从原始海洋中突出的热粘土或沙子促成的。其他理论预测,聚合物的形成是在这种沙子或粘土中的矿物或金属催化剂的帮助下发生的。
研究人员通过将少量富含氨基酸或核酸的溶液滴到热粘土或沙子上测试了这一假设。在许多情况下,单体会在没有催化剂的情况下自发地连接形成聚合物。然而,在这种过程中形成的多肽通常是交联的并且缠绕在一起,并不像现代蛋白质那样。然而,这些分子中有一些具有微弱的催化能力,可能参与了早期地球的某些反应。
原生物在原始上类似于细胞的版本,只是少了“生命”的部分。原生物是包含在膜状结构中的有机分子和其他非生物产物的集合。膜通常由疏水分子聚集体组成,这些聚集体排列成双层结构,类似于质膜的磷脂双层。这些膜结构的形成方式很可能与胶束在水溶液中形成的方式相同。
原生物也已在实验室中以脂质体的形式生成。这些脂质体非常出色,因为围绕内部溶液的疏水双层通常具有选择性渗透性。这种选择性渗透性允许脂质体存储内部溶液,该溶液与周围介质不同。由于这种渗透性和扩散,一些脂质体甚至能够在该疏水双层中维持梯度。已经观察到一些脂质体分裂成更小的物体,尽管没有观察到完美的复制。脂质体被认为是原细胞的可能模型。
最早的细胞出现在还原性大气中(没有氧气),并且最有可能从早期地球上存在的丰富无机燃料中获得能量。
最有可能的是,原始的单细胞生物逐渐获得了从环境中获取化合物的能量的能力,然而,经过多年的进化,生物开始利用能量合成更多的自身前体分子。因此,生物对外部来源的依赖性降低了。
例如,光合作用是通过发展能够转换光能或“固定”CO2成有机化合物的色素来发生的。这种进化变化使植物能够在其环境中更有效地生存。
由于早期地球的大气中几乎没有氧气,因此最早的细胞是厌氧生物,即不依赖氧气存在的生物体。然而,随着光合细菌产生的氧气的增加,大气中的氧气越来越丰富。氧气是一种强氧化剂,对厌氧生物具有致命毒性。在富氧环境中,好氧生物与其厌氧生物相比具有优势,这导致好氧生物在富氧环境中占据主导地位。这种提出的富氧环境发展的时期支持了达尔文的“适者生存”理论,因为好氧生物由于其适应性进化而在环境中繁荣发展。
研究生物进化使生物化学家能够追溯分子解剖学的信息。分子解剖学揭示了进化关系。分子系统发育与传统的宏观系统发育一致,但更精确。从最简单的生物体到最复杂的生物体,分子结构和机制非常相似。当两个基因具有序列相似性时,DNA序列被称为同源的,它们编码的蛋白质被称为同源物。如果同源基因出现在同一种类中,它们被称为旁系同源,它们产生的蛋白质被称为旁系同源物。通常,旁系同源物具有相似的三维结构,但在进化过程中获得不同的功能。另一方面,在不同物种中发现的两个同源基因被称为直系同源,它们的蛋白质产物被称为直系同源物。通常,直系同源物被发现具有相同的功能。因此,可以根据基因组序列猜测基因产物的功能。同源基因之间的差异可以粗略地衡量两个物种在进化过程中分化的程度。序列差异越大,进化史上的分化越早。
查尔斯·达尔文的进化理论指出,在选择压力下,“适者生存”,导致了自然选择的概念。达尔文声称,突变细胞及其后代将在新的环境中生存和繁荣,而野生型(未突变细胞)细胞应该饿死并被淘汰。
1) 物种并非以其现在的形式被创造出来,而是从祖先物种进化而来,这与自然选择无关。
2) 自然选择是进化的机制,它详细说明了生物种群随着时间的推移而发生变化,这是由于具有某些可遗传性状的个体(被认为对其环境有利)能够比其他个体留下更多的后代。
根据查尔斯·达尔文的理论,自然选择是一种现象,通过这种现象,具有有利遗传性状的生物体在生存和繁殖方面变得成功和占主导地位。具有不适合生存的性状的生物体不受自然界的青睐,因此灭绝。这个过程作用于表型,即生物体的表现。生物体获得的表型是其遗传密码或基因型中随机突变的结果。这意味着地球上生命的所有进程都是由无数生物体遗传密码中的随机突变所引导的,而这些突变又受到非随机环境力量的影响,这些力量有利于特定的性状。总的来说,这种机制使得新的生物物种的出现成为可能,这些物种能够适应其环境并繁荣昌盛。总之,自然选择的过程有利于那些在不断变化的环境中具有生存价值的基因。基尔大学的克里斯托弗·埃克斯利赞扬了达尔文关于自然选择作为自然力量的观点。他在他的论文“达尔文、自然选择以及铝和硅的生物必需性”中强调,自然选择在生化进化中与物种形成一样重要,它也定义了铝和硅等元素的必需性。尽管铝和硅在地球上都很丰富,但铝在生物系统中没有必需作用,而硅被认为在生物学上是必需的。硅通过帮助铝以非活性形式从生物系统中选择出来,部分参与了自然选择,这是通过与铝反应形成 HAS(羟基铝硅酸盐)实现的,HAS 保护生物系统免受铝的毒性。这显着降低了铝的生物利用度及其与生物有机化合物的生物反应性,将铝排除在生化进化之外(不参与自然选择)。因此,作为一种自然力量,自然选择定义了硅是必需的,因为硅与生物无机化合物(如金属)反应以保持生物系统的安全,而铝由于缺乏生物利用度,在生物系统中没有必需性。
在 DNA 复制过程中,总是会发生被称为基因突变的错误,这会导致 DNA 序列发生变化。复制过程中的缺陷会导致 DNA 发生变化,产生基因突变。如果基因突变通过生殖细胞遗传下去,那么它们可能是极其有害的,因为突变会导致体内某些功能(如酶促反应)无法发生。然而,即使突变是不可取的,它们也使细胞能够进化并变得更强大。突变使细胞能够做以前无法做到的事情。偶尔,突变会更好地使生物体或细胞在其环境中生存。因此,即使它是一个突变体,如果它试图在一个新的环境中生存,并且数量很多,它将能够比野生型细胞更好地在这个环境中生存,这将导致野生型细胞死亡并变得有限。突变细胞将比其他未突变的“野生型”细胞具有选择优势。最终,野生型细胞会饿死并被淘汰。有时,基因在 DNA 复制过程中会意外地复制自身,这是由于突变造成的。第二个副本是不必要的,对这个基因的突变不会造成损害。因此,这允许基因在保留旧功能的同时获得新功能。当这种情况发生很多次时,第二个副本会发生另一个意外突变,导致测序和由此序列形成的蛋白质发生改变。例如,当己糖激酶基因经历 DNA 复制时,它遇到了一个突变,这造成了它的复制。这个重复基因遇到了另一个错误,导致它形成一个原始基因的副本和一个突变的重复基因的副本,它可能具有不同的能力。在某些情况下,这些突变实际上有助于形成新的基因进化。DNA 由于突变而随着时间的推移发生了很大的进化。适量的突变使 DNA 能够生长并具有变异性,这解释了生物体的多样性。
突变类型
在 DNA 复制过程中发生的突变主要有两类。他们是
- 点突变,其中单个氮碱基发生改变。例如,给定序列:GATTACA。点突变可能导致以下组合:GATAACA。通常,点突变只会改变一个嘧啶碱基与另一个嘧啶碱基,或一个嘌呤与另一个嘌呤。这些突变被称为转换点突变,比它们的对应物(颠换)更常见,颠换是将嘌呤换成嘧啶,反之亦然。转换突变也分为无义突变、错义突变和沉默突变。无义突变涉及意外编码一个终止序列,这会导致形成比最初计划的短得多的蛋白质。错义突变导致编码完全不同的蛋白质,而沉默突变是指蛋白质不受影响的突变。它们可以编码相同或不同的氨基酸,但不会影响整个分子的结构和功能。
- 移码突变,其中添加或删除一个氮碱基会导致蛋白质合成的框架发生移位。例如,如果核糖体一次加载三个氮碱基对的分子,并且它最初计划像这样分离以下内容:/GAT/TAC/A。移码将看起来像这样,带有添加,/AGA/TTA/CA,或者带有删除,/ATT/ACA。
突变的优点
- 突变体可以获得不同的特异性,这些特异性可能更有利或更适合其环境,因此会导致更高的存活率以及该突变在物种基因库中的传播。将一个可育的突变体引入一个物种,可以使遗传差异范围更广,从而增加该物种的活力。
突变的缺点
- DNA 突变可能是有害的甚至致命的,因为可能会发生缺陷,并可能导致酶或其他维持生命所需特异性的缺失或变性,由此产生的缺陷可能导致无法完成适当的功能。例如,许多人类遗传疾病是由 DNA 突变引起的。DNA 突变可能是有害的甚至致命的。在大肠杆菌中,当在 DH5-alpha 细胞的特定菌株中进行点突变时,细菌会变得致命,这会导致生长完全停止并终止细胞。据说复制过程中的许多错误会导致疾病,例如[[Structural Biochemistry/Protein function/Heme group/Hemoglobin/Sickle disease
|镰状细胞性贫血]],其中点突变会导致细胞变形并聚集,从而抑制适当的血流,导致全身剧烈疼痛。紫外线照射或接触致癌物会导致偶尔发生的突变积累并导致癌症。据说体内突变的积累也是导致衰老过程的因素之一。
- 可遗传突变使生物体能够更适应在生态位中生存,并在繁殖方面繁荣,朝着优先选择的性状发展。突变和自然选择的过程是基于达尔文主义进化理论,该理论指出,所有生物体之间的基本相似性源于一开始,第一个单细胞。
基因组的序列,即生物体的完整遗传禀赋,为生物化学家提供了一个极其丰富且不断增长的信息库,这些信息可以用来分析进化关系和完善进化理论。通过进化,结构、过程和机制反映在进化生物体的不断变化的基因组中。因此,比较每个门类的物种的整个基因组正在导致澄清和识别对身体计划和发育中的基本进化变化至关重要的基因。
由于序列的定量比较是可能的,因此将更好地了解进化过程,其中在生物体之间经常发现相似性,而差异将阐明生物体之间多样性的遗传原因。通过识别基因组中编码的途径(酶集),生物化学家和遗传学家可以更多地了解生物体的遗传物质,并仅从基因组序列推断生物体的代谢能力。
数百种细菌、40 种古细菌和众多真核微生物的基因组已知,并且正在不断扩展。
当基因组的序列完全确定,并且每个基因都被分配了一个功能时,分子遗传学家可以根据过程对基因进行分组,从而找到基因组的哪一部分分配给细胞的每个活动。一般来说,生物体越复杂,其基因组中用于调节细胞过程的部分就越大。
大肠杆菌、拟南芥和智人的基因由以下组成:
- 40+% 功能未知
- ~10-4% 转运蛋白
- ~6-2% 编码蛋白质和 RNA
从系统发育树不同分支中获得的有机体完整基因组的新知识提供了对进化的洞察,最终将有利于人类医学。 Image:Full resolution
体外进化,也称为SELEX(通过指数富集进行配体系统进化),是对RNA和DNA核苷酸及其在微观尺度上的自然选择的研究。RNA和DNA功能的研究是通过将核苷酸混合在一起,并在细胞外(而不是细胞内,细胞内称为体内进化)观察它们进行的。将样本置于促进样本中核苷酸之间生存竞争的条件下,这种“竞争”通常与核苷酸特定特性的生存有关。表现出显性生存的核苷酸随后被扩增。
体外进化有几个目的,主要目的是创建核苷酸或蛋白质来执行特定的功能。SELEX还被用于预测核苷酸的进化,假设刺激核苷酸样本可以获得进化的适应性。诱导自适应突变,并将其用于预测核苷酸序列的进化过程,或优化分子(例如蛋白质),以更好地执行特定功能。例如,它可以用来增加抗体的结合亲和力,也可以用来预测耐药性的发展。
进行体外进化研究的方法一直在不断变化。最初,通过在核苷酸水平上诱导来获得诱变。然而,通过聚合酶链式反应 (PCR) 进行核苷酸的单点突变相当困难,因为在某些情况下,只有氨基酸样本的一小部分可以通过这种方法获得。在密码子水平上进行的诱变,例如使用三核苷酸磷酰胺,在实际应用中产生了更多结果,因为它不那么繁琐。可以在 PCR 和密码子水平诱变之间比较功效的例子:在将简并性引入密码子的情况下,会产生 32 个核苷酸序列,而在密码子水平诱变中,只会产生 20 个。当过量的核苷酸呈指数级增长时,使用更高序列组合(PCR)扩增样本在后期会成为负担。
体外进化的实用性至关重要的一点是能够将理想的突变从大量突变群体中分离出来。通常,为了实现朝着特定功能的突变,需要使用基于该功能的快速分离技术。分离和筛选都是用于检测文库的宝贵技术。简单的筛选可能有用,但由于显着且特异性的突变需要大量的突变文库,因此需要更高的通量方法来可靠地找到具有所需突变的个体。因此,选择通常更有用,因为它们可以评估大量突变体。
体外进化最早由索尔·斯皮尔曼 (Sol Spiegelman) 在 1960 年代提出。他从 Qχ² 病毒中提取了 RNA。斯皮尔曼没有使用传统的体内复制方法,而是使用提取的酶,即 RNA Q-b 复制酶(一种病毒 RNA 分子)在试管中合成 RNA。RNA 开始在没有细胞繁殖的情况下进行复制。通过斯皮尔曼的实验,达尔文的适者生存进化理论被证明不仅适用于生物体,也适用于分子。编码成最适合优化条件的 RNA 将继续复制,而竞争力较低的 RNA 将因溶液中有限的核苷酸资源而死亡。因此,在经过几次复制过程后,最初的 4,500 个核苷酸碱基缩减到 218 个短的核苷酸碱基,因为较短的 RNA 链往往复制更快。缩短的 RNA 链被称为斯皮尔曼怪物。
关于体外进化是否能准确地描绘发生在细胞中的进化(体内进化),存在很多争议。然而,罗切斯特大学的米里亚姆·巴洛 (Miriam Barlow) 和巴里·G·霍尔 (Barry G. Hall) 最近的研究表明,体外进化与体内进化有很强的相似性。他们的研究对 TEM-1 β-内酰胺酶基因的体外进化进行了研究,并将它的进化路径与自然发生的进化进行了比较。他们的研究结果发现,体内进化的大部分氨基酸替换也存在于体外进化中。这项研究的结论是,体外进化技术可以可靠地用于预测基因的未来进化。
尽管体外研究可以作为分子水平上的信息性研究,但许多蛋白质和细胞相互依赖地发挥作用。因此,在试管中,许多相互作用和功能与整体生物体不同。
参见主要文章 结构生物化学/内共生理论
原核细胞和真核细胞之间的一个主要区别是真核细胞中存在细胞器。细胞器是存在于真核细胞细胞质中的膜结合“体”,执行特定的功能。
关于这些细胞器是如何获得的一种流行理论是内共生理论。这种理论试图解释线粒体和叶绿体在真核细胞中的出现。该理论背后的核心思想如下:
一个原核细胞吞噬(或被)另一个原核细胞。两个原核细胞都无法破坏或消化对方。结果,较小的细胞在较大的细胞内部存活下来。在线粒体的情况下,较小的原核细胞(一种有氧原核生物)将为宿主细胞提供利用氧气的途径,同时获得宿主细胞吸收的营养物质。这表现出一种共生关系,其中宿主及其共生伙伴都为伙伴关系中每个个体的福祉做出贡献。
“二次内共生”试图解释植物细胞和某些原生生物中线粒体和叶绿体的存在。
支持这一理论的证据包括线粒体中存在双层脂质双层(膜)以及线粒体 DNA 的存在。
- 真核细胞从内共生细菌获得了进行光合作用和氧化磷酸化的能力,而多细胞生物发展出专门的细胞类型,这些细胞类型区分了对生物体生存至关重要的功能。

1. Oparin, Aleksandr Ivanovich. 生命的化学起源. C.C. Thomas, 1964.
2. http://www.sciencemag.org/cgi/content/full/308/5724/962
3. http://www.ux1.eiu.edu/~cfjps/1400/atmos_origin.html
4. Lehninger 和 Nelson 著“Lehninger 生物化学原理”
5. 'Reece 和 Campbell. 生物学,第 7 版. 纽约,2005. ISBN 0-8053-7171-0 '
- Breaker, Ronald. 教程:体外进化. [1] 检索日期为 2008-10-18
- Campbell, Neil A., Jane B. Reece. 生物学. 第七版. 2005. ISBN 0-8053-7171-0
- 预测进化潜力 检索日期为 2008-10-22
- Simon Delagrave, Dennis J. Murphy. 测定和药物开发技术. 2003 年 2 月 1 日,1(增刊 2):187-198. doi:10.1089/154065803321537917。